Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Регуляторные функции гормонов надпочечниковСодержание книги
Поиск на нашем сайте
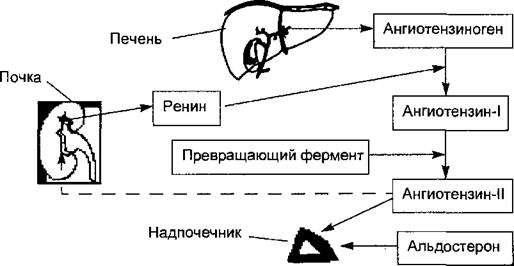
Надпочечники являются парной железой внутренней секреции, морфологически и функционально состоящей из двух разных по эмбриональному происхождению тканей — коркового и мозгового вещества. Кора надпочечников — производное мезодермы, синтезирует три вида гормонов стероидной природы, или кортикостероидов. Мозговое вещество называют также хромаффинной тканью, которая имеет общее с нервной системой происхождение и развивается из эмбриональных симпатических клеток. Кроме надпочечника хромаффинная ткань, образующая катехоламины, имеется в составе симпатического пограничного ствола, брюшном и солнечном сплетении. Кровоснабжение надпочечника осуществляется тремя группами артерий, ч отходящих от диафрагмальной и почечной артерий и аорты, а также не- 4 скольких добавочных артерий. Ветви этих сосудов образуют раздельные корковые и мозговые артерии, дающие обильные капиллярные сети. Венозный отток крови осуществляется по многочисленным венам в систему нижней полой вены и бассейн воротной вены печени. Как в корковом, так < и в мозговом веществе имеется большое количество нервных волокон. Особое значение имеют ветви чревного нерва, несущие пре- и постганглионарные волокна к хромаффинным клеткам мозгового вещества и обеспечивающие поступление к ним регуляторных стимулов. 6.3.1. Гормоны коры надпочечников и их эффекты в организме Кора надпочечников, занимающая по объему 80 % всей железы, состоит h из трех клеточных зон: наружной клубочковой зоны, образующей минералокортикоиды, средней пучковой зоны, образующей глюкокортикоиды, и внутренней сетчатой зоны, в небольшом количестве продуцирующей половые стероиды. Все кортикостероиды образуются из холестерина крови и синтезируемого в самих корковых клетках. При синтезе кортистероидов i образуется порядка 50 различных соединений, однако секретируются в - кровь в физиологических условиях лишь 7—9 из них. ( 6.3.1.1. Регуляция секреции и физиологические эффекты минералокорт икоидов У человека единственным минералокортикоидом, поступающим в кровь, I является альдостерон. Регуляция синтеза и секреции альдостерона осуще- I ствляется преимущественно ангиотензином-П, что дало основание считать альдостерон частью ренин-ангиотензин-альдостероновой системы или регуляторной оси (рис. 6.11), обеспечивающей регуляцию водно-солевого обмена и гемодинамики. Регуляции секреции альдостерона может осуществ-
Рис. 6.11. Ренин-ангиотензин-альдостероновая система. Секреция юкстагломерулярными клетками почек в кровь фермента ренина вызывает отщепление пептида ангиотензина-1 от белка плазмы крови ангиогензиногена, образуемого в печени. В сосудистом русле почек, печени, легких, мозга ангиотензин-1 подвергается воздействию превращающего фермента, вызывающего образование из ангиотензина-1 ангиотензина-2. Ангиотензин-2 стимулирует секрецию альдостерона клубочковой зоной коры надпочечников. Пунктирной стрелкой обозначена отрицательная обратная связь — подавление секреции ренина ангиотензином-2.
ляться и под влиянием собственной адренокортикальной ренин-ангиотен- зиновой системы, что объясняет частое несоответствие уровней активности ренина в плазме крови и секреции альдостерона. Поскольку альдостерон регулирует содержание в крови ионов Na+ и К+, обратная связь в регуляции его секреции реализуется прямым влиянием ионов К+ на клубочковую зону коры надпочечников. В ренин-ангиотензин-альдостероновой системе обратные связи включаются при сдвигах содержания Na+ в моче дистальных канальцев, объема и давления крови. Механизм действия альдостерона, как и всех стероидных гормонов, состоит в прямом влиянии на генетический аппарат ядра клеток со стимуляцией синтеза соответствующих РНК, активации синтеза транспортирующих катионы белков и ферментов, а также повышении проницаемости мембран для аминокислот. Негеномные эффекты гормона реализуются через системы вторичных посредников. Механизм действия альдостерона на клетки почечных канальцев представлен на рис. 6.12. Стимуляция всасывания натрия под влиянием альдостерона происходит не только в нефроне, но и в желудочно-кишечном тракте, протоках желез внешней секреции, желчном пузыре. Негеномные эффекты альдостерона обусловлены стимуляцией мембранного антипорта Na+/H+ в клетках разных типов (гладкие мышцы матки, эпителий дистальных канальцев почек, гладкие мышцы артерий и артериол, клетки крипт кишечника). Эти эффекты обусловлены образованием вторичного посредника диацилглицерола и активацией протеинкиназы С. Повышение уровня внутриклеточного кальция в эндотелиальных и гладкомышечных клетках сосудов под влиянием альдостерона обусловлено активацией вторичного посредника ИФЗ. Альдостерон вызывает в клетках и двукратное повышение уровня цАМФ, модулируя геномные эффекты стероидных гормонов. Быстрые негеномные эффекты альдостерона проявляются и со стороны сердечно-сосудистой системы в виде: повышения сосудистого сопротивления и артери-
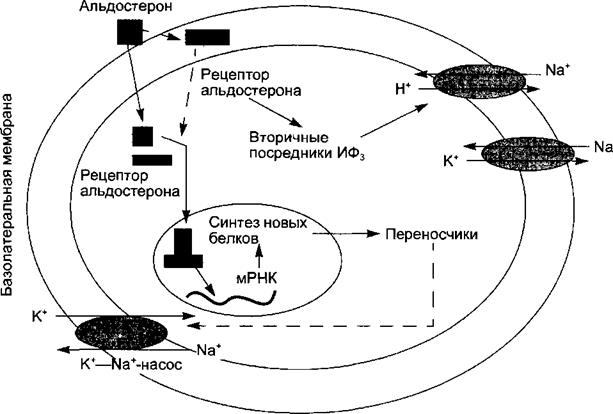
Рис. 6.12. Геномный и внегеномный механизмы действия альдостерона на клетку почечного канальца.
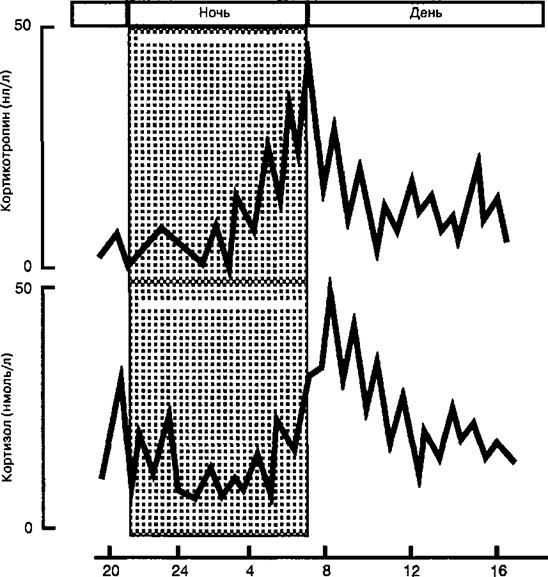
Геномный механизм: проникновение молекулы гормона через мембрану внутрь клетки, связывание с цитоплазматическим рецептором, транспорт в ядро, связывание с ядерным рецептором, активация синтеза белков (Na-транспортирующего белка-переносчика) и Na+-К?-антипорта через люминальную мембрану. Внегеномный механизм: связывание молекулы гормона с мембранным рецептором, образование вторичных посредников (ИФЗ), фосфорилирование и активация Na^-протонного антипорта через люминальную мембрану. ального давления при снижении сердечного выброса, противодействия повышению в гладких мышцах сосудов уровня цАМФ и увеличения чувствительности к прессорным эффектам катехоламинов и ангиотензина-П, что дало основание считать альдостерон циркуляторным гормоном стресса. Альдостерон поддерживает оптимальный водно-солевой обмен между внешней и внутренней средой организма. Одним из главных органов-мишеней гормона являются почки, где альдостерон вызывает усиленную реабсорбцию натрия в дистальных канальцах с его задержкой в организме и повышение экскреции калия с мочой. Под влиянием альдостерона происходит задержка в организме хлоридов и воды, усиленное выведение Н-ио- нов и аммония, увеличивается объем циркулирующей крови, формируется сдвиг кислотно-основного состояния в сторону алкалоза. Действуя на клетки сосудов и тканей, гормон способствует транспорту натрия и воды во внутриклеточное пространство. Минералокортикоиды являются жизненно важными гормонами, гибель организма после удаления надпочечников можно предотвратить, вводя гормоны извне. Минералокортикоиды усиливают воспаление и реакции иммунной системы. Избыточная их продукция ведет к задержке в организме натрия и воды, отекам и повышению артериального давления, потере калия и водородных ионов, вследствие чего возникают нарушения возбудимости нервной системы и миокарда. Недостаток альдостерона у человека сопровождается уменьшением объема крови, гиперкалиемией, гипотензией, угнетением возбудимости нервной системы. 6.3.1.2. Регуляция секреции и физиологические эффекты глюкокортикоидов Клетки пучковой зоны секретируют в кровь у здорового человека два основных глюкокортикоида: кортизол и кортикостерон, причем кортизола примерно в 10 раз больше. Секреция глюкокортикоидов регулируется кортикотропином аденогипофиза. Избыток кортизола в крови по механизму обратной связи угнетает секрецию кортиколиберина в гипоталамусе и кортикотропина в гипофизе. Секреция глюкокортикоидов происходит непрерывно с отчетливой суточной ритмикой, повторяющей ритмику секреции кортикотропина: максимальные уровни гормона в крови у человека отмечаются в утренние часы, а минимальные — вечером и ночью (рис. 6.13). Поступающие в кровь гормоны транспортируются к тканям в свободной и связанной (до 95 %) с а2-глобулином плазмы (транскортин) формах. Механизм действия глюкокортикоидов на клетки-мишени представлен на рис. 6.14.
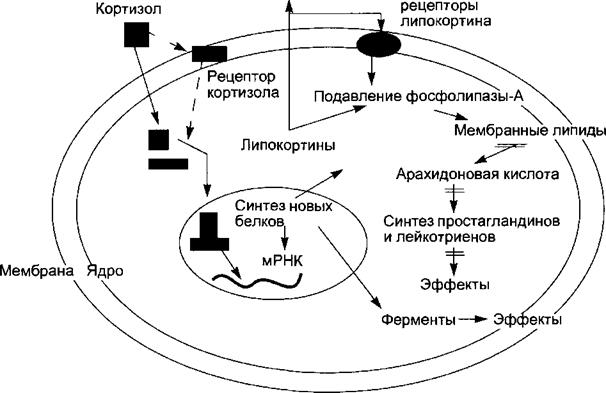
Благодаря растворимости в липидах кортизол проникает через мембрану клетки-мишени и взаимодействует с цитоплазматическим рецептором, образуя лиганд-рецепторный комплекс, что обеспечивает транспорт молекулы гормона в ядро, где кортизол связывается с ядерным рецептором, активируя синтез новых белков и ферментов, тем самым обеспечивая метаболические эффекты. Молекула кортизола может образовывать лиганд-рецепторный комплекс и с мембранными рецепторами. Хотя роль этого процесса в реализации эффектов гормона еще изучается, тем не менее известны быстрые негеномные эффекты гормона на возбудимость нервных клеток, связанные с изменением ионного трансмембранного транспорта, обусловливающие изменение поведения. Среди синтезированных под влиянием кортизола белков в клетке-мишени важнейшую роль для реализации опосредованных эффектов гормона играет семейство липокортинов. Последние, выходя из клетки, связываются со специфическими липокортиновыми рецепторами клеточной мембраны (аутокринный путь влияния), что вызывает подавление активности фосфолипазы-А. Липокортины способны ингибировать фосфолипазу-А и непосредственно, следствием чего является подавление синтеза в клетках простагландинов и лейкотриенов, ослабление их метаболических и регуляторных эффектов. Уменьшение проницаемости клеточных мембран и противовоспалительный эффект кортизола обусловлены ингибированием синтеза лейкотриенов. Глюкокортикоиды прямо или опосредованно регулируют практически все виды обмена веществ и физиологические функции. Метаболические эффекты глюкокортикоидов проявляются, прежде всего, со стороны углеводного, белкового и жирового обменов. Обобщенно эти сдвиги можно свести к распаду белков и липидов в тканях, после чего метаболиты поступают в печень, где из них синтезируется глюкоза, использующаяся как источник энергии. Эффекты на углеводный обмен в целом противоположны инсулину, поэтому глюкокортикоиды называют контринсулярными гормонами. Гипергликемия под влиянием гормонов возникает за счет усиленного образования глюкозы в печени из аминокислот — глюконеогенеза и подавления утилизации ее тканями. Гипергликемия является причиной активации секреции инсулина. Чувствительность тканей к инсулину глюкокор-
Рис. 6.13. Суточный ритм секреции кортикотропина и кортизола. Максимум концентрации гормонов в крови приходится на утренние часы, при этом прирост уровня кортикотропина опережает рост содержания кортизола.
тикоиды снижают, а контр инсулярные метаболические эффекты могут вести к развитию стероидного сахарного диабета. На белковый обмен гормоны оказывают катаболический и антианаболический эффекты, приводя к отрицательному азотистому балансу. Распад белка происходит в мышечной, соединительной и костной тканях, снижается уровень альбумина в крови, уменьшается проницаемость клеточных мембран для аминокислот. Однако синтез некоторых белков в печени, например а2-глобулинов, глюкокортикоиды повышают. Со стороны жирового обмена имеют место липолитический эффект в тканях, гиперлипидемия и гиперхолестеринемия, активация кетогенеза в печени, угнетение липогенеза в печени, стимуляция липогенеза и перераспределения жира в жировой ткани центральной оси туловища и лица, стимуляция аппетита и потребления жира Влияние глюкокортикоидов на реактивность тканей проявляется не только в виде подавления чувствительности к инсулину, но и в повышении чувствительности адренорецепторов к катехоламинам. Глюкокортикоиды вызывают снижение в крови количества лимфоцитов, эозинофилов Эффекты Эффект через
Рис. 6.14. Схема механизма действия кортизола на клетку-мишень. Проникая внутрь клетки через мембрану, молекула гормона последовательно взаимодействет с цитозольным, а затем с ядерным рецептором. Следствие геномного влияния — активация синтеза новых белков, в том числе являющихся внутриклеточными ферментами, что вызывает изменения обмена веществ. К числу синтезируемых под влиянием кортизола белков относятся липокортины. Последние либо выводятся из клетки и взаимодействуют со специфическими для них мембранными рецепторами, либо действуют внутриклеточно. Основной эффект липо- кортинов — ингибирование мембранного фермента фосфолипазы-А и образования из арахидоновой кислоты простагландинов и лейкотриенов.
и базофилов, повышение чувствительности сенсорных структур и возбудимости нервной системы, обеспечение оптимальной симпатической регуляции сердечно-сосудистой системы. Почечные эффекты глюкокортикоидов состоят в стимуляции диуреза путем снижения реабсорбции воды и повышении клубочковой фильтрации; подобно минералокортикоидам, они могут вызывать задержку натрия при потере калия. Глюкокортикоиды повышают синтез ангиотензиногена в печени и тем самым способствуют большему образованию в крови ангиотензина-П и секреции альдостерона, увеличивают синтез катехоламинов в мозговом веществе надпочечников. Гормоны повышают устойчивость организма к действию чрезмерных раздражителей, подавляют сосудистую проницаемость и воспаление (поэтому их называют адаптивными и противовоспалительными), из-за катаболизма белка в лимфоидной ткани и угнетения иммунных реакций они оказывают антиаллергические эффекты. Физиологическое влияние кортизола на иммунокомпетентные клетки носит защитный, регуляторный характер (рис. 6.15).
Избыток гормонов в крови вызывает активацию желудочной секреции, выделения НС1, уменьшение числа мукоцитов и продукции слизи, что способствует возникновению язвы желудка — ульцерогенный эффект.
ФНО и другие токсичные соединения Иммунологический стимул Рис. 6.15. Физиологическое значение влияния кортизола на иммунокомпетентные клетки (на примере макрофага). Кортизол тормозит выделение макрофагом фактора некроза опухолей (ФНО) и других токсичных для клеток организма макрофагальных цитокинов. Под влиянием иммунологического стимула макрофаг выделяет кортикотропин, что усиливает продукцию кортизола надпочечниками и ослабляет выделение макрофагом цитотоксические соединений. Кроме того выделяемые макрофагом интерлейкины, в частности интерлейкин-1, стимулируют гипоталамо-адено- гипофизарно-надпочечниковую ось регуляции, что также повышает продукцию кортизола, снижающего цитотоксические эффекты макрофага. Повышенная секреция глюкокортикоидов, не связанная с описанной выше усиленной секрецией кортикотропина, вызывает развитие синдрома Иценко—Кушинга. Его основные проявления близки болезни Иценко-Ку- шинга, однако, благодаря механизму обратной связи, избыток глюкокортикоидов угнетает секрецию кортикотропина и усиленную пигментацию. Среди проявлений повышенной секреции гормонов: антиростовые эффекты (подавление действия кальцитриола и митоза фибробластов, деградация коллагена вплоть до остеопороза; подавление заживления ран из-за угнетения пролиферации фибробластов и редукции соединительной ткани; подавление секреции соматотропина и синтеза ИФР-1; мышечная атрофия и слабость), склонность к сахарному диабету, гипертензия, нарушения половых функций, лимфопения, пептические язвы желудка, изменения психики, предрасположенность к инфекциям. Дефицит глюкокортикоидов вызывает гипогликемию, снижение чувствительности сердечно-сосудистой системы к симпатической регуляции, замедление сердечного ритма, гипотензию, нейтропению, эозинофилию и лимфоцитоз, снижение сопротивляемости организма инфекциям. 6.3.1.3. Регуляция секреции и физиологические эффекты половых стероидов коры надпочечников Клетками сетчатой зоны у человека секретируются в кровь мужские половые гормоны или андрогеньг. андростендион, дегидроэпиандростерон и существенно меньше — 11-бета-гидроксиандростендион. Наиболее высокий уровень этих гормонов отмечается в 6 часов утра, а наиболее низкий — в 19 часов. Секреция андрогенов регулируется кортикотропином гипофиза. Андрогены надпочечника стимулируют окостенение эпифизарных хрящей, повышают синтез белка (анаболический эффект) в коже, мышечной и костной ткани, а также формируют половое поведение у женщин. Гормоны являются предшественниками основного андрогена семенников — тестостерона и могут превращаться в него при метаболизме в тканях-мишенях. Андрогены надпочечников способствуют развитию оволосения по мужскому типу, а их избыток у женщин — к вирилизации, т. е. появлению мужских черт. Ранний избыток андрогенов ведет к преждевременному проявлению вторичных половых признаков у мальчиков и вирилизации женских половых органов. Нарушения секреции андрогенов коры надпочечников получили название адреногенитальных синдромов. 6.3.2. Гормоны мозгового вещества надпочечников и их эффекты в организме Мозговое вещество надпочечников содержит хромаффинные клетки, названные так из-за избирательной окраски хромом. По происхождению и функции они являются постганглионарными нейронами симпатической нервной системы, однако, в отличие от типичных нейронов, клетки надпочечников: 1) синтезируют больше адреналина, а не норадреналина (отношение у человека между ними 6:1); 2) накапливая секрет в гранулах, после поступления нервного стимула они немедленно выбрасывают гормоны в кровь. Регуляция секреции гормонов мозгового вещества надпочечников осуществляется благодаря наличию гипоталамо-симпатоадреналовой оси, при этом симпатические нервы стимулируют хромаффинные клетки через холинорецепторы, выделяя медиатор ацетилхолин. Хромаффинные клетки являются частью общей системы нейроэндокринных клеток организма, или APUD-системы (Amine and amine Precursors Uptake and Decarboxylation), т. e. системы поглощения и декарбоксилирования аминов и их предшественников. К этой системе относятся нейросекреторные клетки гипоталамуса, клетки желудочно-кишечного тракта (энтериноциты), продуцирующие кишечные гормоны, клетки островков Лангерганса поджелудочной железы и К-клетки щитовидной железы. Гормоны мозгового вещества — катехоламины — образуются из аминокислоты тирозина поэтапно: тирозин—ДОФА—дофамин-норадреналин— адреналин. Хотя надпочечник и секретирует значительно больше адреналина, тем не менее в состоянии покоя в крови содержится в четыре раза больше норадреналина, так как он поступает в кровь и из симпатических окончаний. Секреция катехоламинов в кровь хромаффинными клетками осуществляется с обязательным участием Са2+, кальмодулина и особого белка синексина, обеспечивающего агрегацию отдельных гранул и их связь с фосфолипидами мембраны клетки. Катехоламины называют гормонами срочного приспособления к действию сверхпороговых раздражителей среды. Физиологические эффекты катехоламинов обусловлены различиями в адренорецепторах (альфа и бета) клеточных мембран, при этом адреналин обладает большим сродством к бета-адренорецепторам, а норадреналин — к альфа. Чувствительность адренорецепторов к адреналину увеличивают гормоны щитовидной железы и глюкокортикоиды. Основные функциональные эффекты адреналина проявляются в виде: 1) учащения и усиления сердечных сокращений, 2) сужения сосудов кожи и органов брюшной полости, 3) повышения теплообразования в тканях, 4) ослабления сокращений желудка и кишечника, 5) расслабления бронхиальной мускулатуры, 6) стимуляции секреции ренина почкой, 7) уменьшения образования мочи, 8) повышения возбудимости нервной системы, скорости рефлекторных процессов и эффективности приспособительных реакций. Адреналин вызывает мощные метаболические эффекты в виде усиленного расщепления гликогена в печени и мышцах из-за активации фосфорилазы, а также подавление синтеза гликогена, угнетение потребления глюкозы тканями, что в целом ведет к гипергликемии. Адреналин вызывает активацию распада жира, мобилизацию в кровь жирных кислот и их окисление. Все эти эффекты противоположны действию инсулина, поэтому адреналин называют контринсулярным гормоном. Адреналин усиливает окислительные процессы в тканях и повышает потребление ими кислорода. Таким образом, как кортикостероиды, так и катехоламины обеспечивают активацию приспособительных защитных реакций организма и их энергоснабжение, повышая устойчивость организма к неблагоприятным влияниям среды. В мозговом веществе надпочечников, кроме катехоламинов, образуется и пептидный гормон адреномедуллин. Кроме мозгового вещества надпочечников и плазмы крови он выявлен в тканях легких, почек и сердца, а также эндотелиальных клетках сосудов. Этот пептид состоит у человека из 52 аминокислот. Основное действие гормона заключается в мощном сосудорасширяющем эффекте, в связи с чем его называют гипотензивным пептидом. Второй физиологический эффект гормона заключается в подавлении продукции альдостерона клетками клубочковой зоны коры надпочечников. При этом пептид подавляет не только базальный, фоновый уровень образования гормона, но и его секрецию, стимулированную высоким уровнем калия в плазме крови или действием ангиотензина-И.
|
||||||||||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2020-12-09; просмотров: 206; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.144.104.118 (0.017 с.) |