Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Функции лимбической системы мозгаСодержание книги
Поиск на нашем сайте
Лимбическая система участвует в формировании целенаправленного поведения: она определяет возникновение мотиваций (побуждений к действию) и эмоций, способствует научению и образованию следов долговременной памяти. Структуры мозга, входящие в эту систему, объединены пучками нервных волокон в кольцо, охватывающее верхнюю часть ствола (рис. 4.29). Мамиллярные тела, миндалины, гиппокамп, свод и перегородка соединены несколькими пучками аксонов так, что нервные импульсы распространяются от одной лимбической структуры к другой. Гипоталамус соединен с гиппокампом и перегородкой волокнами свода, а терминальной полоской (stria terminalis) и вентральным амигдалофугальным путем — с миндалиной. В лимбическую систему включают также обонятельный мозг, гипоталамус и соседние области древней и старой коры мозга: прилегающие к гиппокампу грушевидные доли и поясные извилины, которые занимают медиальную поверхность больших полушарий непосредственно над мозолистым телом и, как пояс, огибают таламус.
Продольный — дорсальный К медиальной части переднего мозга Входящие в лимбическую систему структуры имеют двусторонние связи с вентральной покрышкой среднего мозга и благодаря этому способны изменять характер управления вегетативными и двигательными реакциями, контролируемыми ретикулярной формацией, а ретикулярная формация, со своей стороны, регулирует активность лимбической системы. С помощью передних ядер таламуса лимбическая система имеет связь с лобной корой, контролирующей активность лимбической системы. Гиппокамп и миндалины, расположенные в височных долях, непосредственно сообщаются с височной корой, через которую получают переработанную в ней информацию от зрительной, слуховой и соматосенсорной коры.
Ассоциативные области коры, функционально взаимодействующие с лимбической системой, относят к лимбической коре и включают в нее медиальные и вентральные поверхности лобных долей, часть медиальной поверхности затылочных долей, а также обращенную кпереди поверхность височных долей. Лимбическая кора совместно с лимбической системой организует мотивационные и эмоциональные аспекты поведения, участвует в образовании долговременной памяти и формировании научения. Она интегрирует информацию, полученную одновременно от лимбической системы и регионов вторичной сенсорной коры, после чего оказывает влияние на ассоциативные области коры, создающие план предстоящих действий. Взаимодействие лимбических структур и коры является физиологической основой для формирования субъективных переживаний эмоций, а связи между лимбической системой и стволом служат для возникновения вегетативных и двигательных реакций, являющихся еще одним компонентом эмоций. Во время проведения стереотаксических операций некоторым категориям больных была установлена зависимость между искусственной электрической стимуляцией различных лимбических структур и субъективным переживанием приятных ощущений (радость, комфорт, покой) или, напротив, неприятных (тревога, беспокойство, уныние). К позитивным эмоцио- генным зонам принадлежат латеральный и перивентрикулярный гипоталамус, перегородка, покрышка среднего мозга. Негативных эмоциогенных зон в мозгу гораздо меньше, чем позитивных, а многие структуры (и в том числе, по-видимому, гиппокамп) нейтральны — их стимуляция не сопровождается приятным или неприятным ощущением. В мозгу человека больше областей, стимуляция которых вызывает приятные ощущения. Естественное возбуждение эмоциогенных структур и соответствующее ему возникновение эмоций происходит в результате модуляции их деятельности дофаминергическими нейронами вентральной покрышки среднего мозга. Волокна дофаминергических нейронов идут в составе медиального пучка переднего мозга и образуют диффузные проекции на перегородку, обонятельные луковицы, миндалины, прилегающее ядро переднего мозга и фронтальную лимбическую кору. Субъективные ощущения, переживаемые человеком как эмоции, фиксируются в памяти, и поэтому он стремится к повторению своих действий, если они вели к переживанию положительных эмоций, и избегает совершать действия, связанные с возникновением отрицательных эмоций.
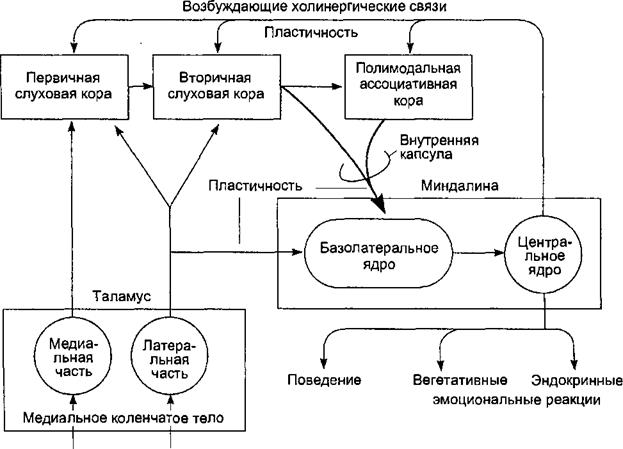
4.8.1. Функции миндалин Миндалевидный комплекс ядер находится в глубине передней части височной доли, его базолатеральные ядра получают афферентные проекции из двух источников: 1) от сенсорных ядер таламуса и 2) от височной и орбитофронтальной коры (рис. 4.30). Эмоциогенные стимулы, действующие на организм, активирует миндалины с помощью короткого прямого пути передачи сигналов от сенсорных ядер таламуса, лишь после этого к ним поступает информация о том же раздражителе, предварительно переработанная в коре. Миндалины сопоставляют сенсорную информацию, поступившую от таламуса, с информацией, пришедшей из коры, чтобы выделить сигналы опасности. Сличение информации происходит в базолатеральном ядре миндалины, а затем в ее центральном ядре формируется эфферентный ответ. Нейроны центрального ядра образуют прямые и непрямые проекции на другие лимбические структуры, гипоталамус, ствол, поясную извилину, орбитофронтальную кору и ассоциативные области коры. В угрожающей ситуации нейроны базолатеральных ядер миндалин немедленно активируются, что происходит, например, при вызывающем боль действии электрического тока. Если пришедшая чуть позже информация из коры подтверждает наличие опасности, нейроны центрального ядра генерируют эфферентный ответ миндалин. Миндалины сохраняют следы долговременной памяти об эмоциональном значении стимула, и поэтому после их удаления или искусственной блокады синапсов исчезают не только врожденные реакции на угрожающие стимулы, но и приобретенная ранее способность оценивать ситуацию как угрожающую по косвенным признакам. После двустороннего удаления части височных долей, включая минда-
Лемнисковый Экстра путь слуховой лемнисковый системы путь Рис. 4.30. Схема вовлечения миндалин мозга в ответную реакцию на угрожающие слуховые раздражители.
Миндалины получают прежде всего афферентную информацию от сенсорных ядер таламуса, а затем к ним поступают сигналы о том же раздражителе, но после переработки информации о нем в коре. В базолатеральном ядре происходит сличение информации, поступившей из обоих источников, на этой основе нейроны центрального ядра создают выходные сигналы. лины и гиппокамп, у обезьян возникает комплекс поведенческих расстройств, определяемый как синдром Клювера—Бъюси и характеризующийся прожорливостью, или гиперфагией, гиперсексуальностью и исчезновением чувства страха. Все проявления этого синдрома обусловлены нарушением способности оценивать значение информации, поступающей из окружающей среды, и связывать ее со своим собственным состоянием. После двустороннего удаления только миндалин нарушается социальное поведение обезьян, что проявляется замкнутостью, покорностью, постоянным стремлением к уединению. Миндалины участвуют и в возникновении положительных эмоций, например удовольствия, связанного с определенной пищей (ее видом, вкусом и запахом), они определяют предпочтение той или иной жидкости для питья, определенного места обитания, выбор полового партнера. Некоторые нейроны миндалин чувствительны к действию половых гормонов, и, благодаря своим связям с обонятельными луковицами, способны изменять активность под влиянием феромонов — веществ, выделяемых особями противоположного пола во внешнюю среду. При возникновении конкурирую-
Эффект, обусловленный ЖЖ’ йЖ: Симпатическая активаций
щих мотиваций миндалины обеспечивают выбор доминирующей потребности, определяя ее на основе зафиксированной предшествующим опытом эмоциональной оценки событий. В результате такой оценки миндалины могут переключать цели поведения с второстепенных на наиболее значимую. После удаления миндалин такое переключение становится почти невозможным. Выходная активность миндалин адресована лимбической коре, а также гипоталамусу и стволу мозга. Активация эфферентного пути от миндалин к орбитофронтальной коре и поясной извилине приводит к осознанию переживаемых эмоций. Одновременная передача информации от миндалин к латеральному гипоталамусу и стволу мозга активирует центры регуляции вегетативных и эндокринных реакций, сопровождающих, например, возникновение страха: увеличение частоты сокращений сердца, учащение дыхания и т. д. (рис. 4.31). Эти периферические реакции способствуют мобилизации энергии для реакций борьбы или бегства.
4.8.2. Функции гиппокампа Гиппокамп расположен в медиальной части височной доли, афферентную информацию он получает от близлежащей энторинальной области коры, в которой происходит переключение сигналов, приходящих из нескольких источников. Это, во-первых, сенсорные и ассоциативные области коры, в которых перерабатывается информация обо всех действующих сенсорных стимулах. Во-вторых, это мотивационные структуры мозга, прежде всего гипоталамус, перегородка и миндалины. В третьих, это фронтальная ассоциативная кора, определяющая стратегию поведения. В четвертых, ретикулярная формация, регулирующая активность корковых нейронов и связанная с гиппокампом волокнами свода. Функция гиппокампа состоит в регуляции распределения всех поступающих информационных потоков, классификации таких потоков и создании условий для образования энграмм (следов) памяти. Двустороннее удаление гиппокампа у человека приводит к антероградной амнезии — утрате способности образовывать новую долговременную память при сохранении памяти о событиях, предшествовавших операции. Двустороннее удаление гиппокампа у экспериментальных животных не позволяет им вырабатывать новые поведенческие навыки. После кратковременной стимуляции входящих в гиппокамп волокон обонятельного тракта его пирамидные нейроны начинают разряжаться необычно долго, на протяжении нескольких суток и даже недель, что было названо долговременной потенциацией. Она возникает в глутаматергических синапсах при участии NMDA-рецепторов, связанных с мембранными каналами для ионов кальция. Ток этих ионов в постсинаптический нейрон вызывает активацию вторичных посредников и последующее изменение метаболической активности не только постсинаптического, но и пресинаптического нейрона. При повторяющейся активации одних и тех же афферентных входов, конвергирующих к гиппокампу, в таких синапсах происходит структурная перестройка, которая повышает эффективность синаптической передачи и является основой долговременной памяти. В то же время гиппокамп не является единственным регионом мозга, где происходит долговременная потенциация, как не является и единственным местом хранения следов о предшествующем индивидуальном опыте. Память о прежних событиях сохраняется и после удаления гиппокампа, а некоторые разновидности долговременной памяти могут образоваться и без него. 4.9. Функции базальных ганглиев (стриопаллидарная система) К базальным ганглиям относят хвостатое ядро и скорлупу, вместе образующие полосатое тело (corpus striatum, а также neostriatum), бледный шар (globus pallidus, pallidum) и субталамическое ядро (рис. 4.32). С базальными ганглиями функционально связана черная субстанция среднего мозга, состоящая из ретикулярной части, сходной по происхождению с внутренним сегментом бледного шара, и компактной части, образующей дофаминергическую проекцию на полосатое тело. Наиболее изученная функция базальных ганглиев заключается в их участии в планировании и инициации произвольных движений. Наряду с этим базальные ганглии участвуют в познавательной деятельности мозга, а также в формировании эмоций.
4.9.1. Взаимодействие базальных ганглиев с другими структурами мозга Афферентная информация поступает к базальным ганглиям из нескольких регионов коры\ префронтальной ассоциативной коры, первичной и вторичной моторной коры, соматосенсорной коры, теменных и височных областей ассоциативной коры. Все афферентные волокна оканчиваются на нейронах полосатого тела, которые объединяются в функциональные модули, однотипные по способу переработки информации. Соматотопически организованные волокна нейронов моторных и сенсорных регионов коры направляются к соответствующим им модулям, в результате пучки волокон распределяются в виде отдельных полосок, из-за которых полосатое тело

Базальные ганглии включают хвостатое ядро и скорлупу, вместе образующие полосатое тело, бледный шар и субталамическое ядро; с базальными ганглиями взаимодействует черная субстанция среднего мозга. Базальные ганглии взаимодействуют с корой при планировании и инициации произвольных движений, при этом образуются две цепи нейронных переключений: 1) кора—полосатое тело—бледный шар—таламус—кора; 2) кора—полосатое тело—бледный шар—таламус—полосатое тело. получило свое название. Окончания волокон кортикальных нейронов образуют с нейронами полосатого тела возбуждающие глутаматергические синапсы. В базальных ганглиях входящие сигналы распределяется независимо друг от друга по параллельным путям переработки информации. Эти пути образуют несколько самостоятельных функциональных кругов, включающих в себя помимо базальных ганглиев разные регионы коры. В зависимости от того, какая область коры включена в функциональный круг, определяется и специфическая роль каждой структуры базальных ганглиев. Скорлупа участвует преимущественно в координации движений тела, одни регионы хвостатого ядра необходимы для регуляции движений глаз, другие — для формирования побуждений к конкретным действиям, выбора стратегии поведения или осуществления когнитивных (познавательных) функций. Вентральные отделы полосатого тела участвуют в формировании эмоций. Скелетомоторный круг нейронных переключений используется при планировании и осуществлении контроля произвольной двигательной активности человека. Вначале активируются премоторная и моторная лобные области коры, которые возбуждают топографически соответствующие им модули полосатого тела. Выходная активность полосатого тела распределяется между внутренней частью бледного шара и ретикулярной частью черной субстанции среднего мозга, для которых характерна спонтанная фоновая активность. Под влиянием преобладающих в полосатом теле эфферентных тормозных ГАМКергических нейронов эта активность уменьшается. Значительно меньшая часть выходных нейронов полосатого тела использует в качестве медиатора вещество Р, оказывающее возбуждающее действие. Одновременно с этим многие нейроны полосатого тела синтезируют энкефалин, действующий в качестве нейромодулятора, и суммарный итог действия полосатого тела на бледный шар и черную субстанцию оказывается тормозно-модулирующим. Находящаяся под контролем полосатого тела выходная активность нейронов бледного шара и черной субстанции адресуется клеткам моторных вентролатеральных ядер таламуса. Эти нейроны образуют проекцию на премоторные области коры, замыкая большой круг нейронных переключений: премоторные и моторные области коры—полосатое тело—бледный шар или ретикулярная часть черной субстанции—таламус—премоторная кора. Нейроны моторных ядер таламуса кроме премоторной коры образуют проекцию на клетки полосатого тела: так замыкается малый круг переключений между полосатым телом — бледным шаром или черной субстанцией — таламусом — и вновь полосатым телом. Субталамическое ядро получает топографически организованные сигналы сигналы от премоторной и моторной коры, в соответствии с которыми оно регулирует выходную активность бледного шара. Распространение возбуждения по этим кругам нейронных переключений происходит до того момента, пока базальные ганглии не выберут из предложенного корой ряда моторных программ наиболее приемлемую для конкретной ситуации, а затем инициируют ее реализацию. Конечным результатом такого взаимодействия нейронов является создание в моторной коре двигательных программ, предназначенных для исполнения двигательными центрами ствола и нейронами спинного мозга. Окуломоторный круг, образующийся при участии базальных ганглиев, используется для регуляции произвольных движений глаз, связанных с привлечением внимания к одному из объектов, находящихся в зрительном поле. Этот круг объединяет афферентную информацию от заднетеменной области коры (поле 7) и небольшого по площади фронтального глазного поля (поле 8 по Бродману). В поле 7 коры создается нейронная карта окружающего зрительного пространства, а поле 8 необходимо для регуляции движений глаз, фиксирующих определенную часть этого пространства. Входные сигналы от обоих полей поступают к соответствующим им модулям хвостатого ядра, которое в зависимости от характера сигналов определяет активность нейронов бледного шара и ретикулярной части черной субстанции. Выходная активность последних адресована тем нейронам таламуса, которые связаны с фронтальным глазным полем и влияют на активность его нейронов. Активность одиночных нейронов базальных ганглиев повышается еще до начала произвольных движений контралатеральных конечностей, что указывает на их участие в планировании и формировании начальной стадии произвольных движений. Базальные ганглии не имеют непосредственного выхода к моторным ядрам ствола и спинного мозга и поэтому не влияют на исполнение уже начавшихся движений, а их электрическая активность не коррелирует с активностью и силой сокращения отдельных мышц. Наличие в составе нейронных кругов переключения возбуждающих и тормозных нейронов позволяет сбалансировать выходную активность двигательных систем так, чтобы обеспечить необходимую плавность движений. Базальные ганглии облегчают одни и угнетают другие движения, что способствует чередованию тонических и фазных компонентов движения, т. е. сначала формированию позы, требующейся для совершения конкретного движения, а затем и самого движения. Одним из источников афферентной информации, доставляемой к базальным ганглиям, являются нейроны лобных ассоциативных областей коры: префронтальной и орбитофронтальной, причастных к программированию поведения и эмоционально окрашенному прогнозу возможных результатов предстоящих действий. Их глутаматергические аксоны оканчиваются на нейронах полосатого тела, далее формируется большой круг нейронных переключений, в образовании которого участвуют бледный шар и ретикулярная часть черной субстанции, а затем ассоциативные ядра таламуса, замыкающие круг в ассоциативной коре. С этим кругом нейронных переключений связаны формирование мотиваций поведения, прогнозирование его результатов, познавательная деятельность и возникновение эмоций. 4.9.2. Модуляции нейронных переключений в базальных ганглиях Помимо кортикостриатной проекции полосатое тело получает афферентную информацию от нескольких ядер таламуса, черной субстанции среднего мозга, миндалин, ядер шва продолговатого мозга и нейронов голубого пятна. В результате в каждом модуле полосатого тела создается сложная сеть переключений из возбуждающих глутаматергичеких окончаний корковых нейронов, модулирующих дофаминергических окончаний нейронов компактной части черной субстанции, серотонинергических окончаний нейронов ядер шва и норадренергических окончаний нейронов голубого пятна. В переработке информации участвуют также локальные интернейроны полосатого тела, синтезирующие ацетилхолин и несколько пептидных нейротрансмиттеров. Дофаминергические нейроны компактной дорсальной части ретикулярной формации образуют диффузную нигростриатную проекцию на нейроны полосатого тела. Эта проекция топографически не организована и предназначена для модуляции кортикостриатных глутаматергических переключений. При дегенерации дофаминергических нейронов черной субстанции нарушается необходимый баланс активности нейронов полосатого тела, что приводит к характерным двигательным нарушениям. Дисфункция, вызванная потерей дофаминергических нейронов, проявляется в виде болезни Паркинсона и характеризуется мышечной ригидностью (значительным повышением тонуса мышц), резким ограничением сопутствующих движений, или акинезией, а также появлением крупноразмашистого тремора ~ дрожания конечностей во время покоя. Указанные нарушения двигательной функции возникают в связи с уменьшением влияния дофаминергических нейронов и изменением баланса активности в пользу остальных нейронов полосатого тела. Если же этот баланс нарушается вслед Модулирующее влияние на нейронные переключения в базальных ганглиях оказывают не только дофамин, но и другие биогенные амины (серотонин, норадреналин), а также многочисленные пептидные нейротрансмиттеры. Столь сложный контроль деятельности базальных ганглиев отражает важность выполняемых ими функций.
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2020-12-09; просмотров: 128; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.138.143.25 (0.016 с.) |