Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Пищеварение в ротовой полости и функция глотанияСодержание книги
Поиск на нашем сайте
/Z.4.Z. Ротовая полость Ротовая полость является начальным отделом пищеварительного тракта, где осуществляется анализ вкусовых свойств веществ и разделение их на пищевые и отвергаемые, защита пищеварительного тракта от попадания некачественных пищевых веществ и экзогенной микрофлоры, измельчение, смачивание слюной пищи, начальный гидролиз углеводов и формирование пищевого комка. Кроме того происходит раздражение механо-, хе- мо-, терморецепторов, вызывающее рефлекторное возбуждение деятельности слюнных желез, желез желудка, поджелудочной железы, печени, желез двенадцатиперстной кишки. Защита организма от патогенной микрофлоры, попадающей в ротовую полость, осуществляется благодаря наличию в слюне бактерицидного вещества лизоцима (муромидазы), антивирусному действию нуклеазы слюны, способности иммуноглобулина А слюны связывать экзотоксины, а также в результате фагоцитоза лейкоцитов, содержащихся в слюне. Пища находится в ротовой полости 16—18 с, и за это время слюна смачивает сухие вещества пищи, растворяет растворимые и обволакивает твердые ее частички, нейтрализует раздражающие жидкости или уменьшает их концентрацию, облегчает удаление несъедобных (отвергаемых) веществ, смывая их со слизистой оболочки ротовой полости. 11.4.2. Слюноотделение У человека имеется три пары больших слюнных желез {околоушные, подъязычные, подчелюстные) и большое количество мелких желез, локализованных в слизистой оболочке рта. Слюнные железы состоят из слизистых и серозных клеток. Первые выделяют мукоидный секрет густой консистенции, вторые — жидкий, серозный или белковый. Околоушные слюнные железы содержат только серозные клетки. Такие же клетки находятся и на боковых поверхностях языка. Подчелюстные и подъязычные содержат как серозные, так и слизистые клетки. Подобные железы расположены и в слизистой оболочке губ, щек, на кончике языка. Подъязычные и мелкие железы слизистой оболочки выделяют секрет постоянно, а околоушные и подчелюстные — при их стимуляции. Ежедневно у человека продуцируется от 0,5 до 2,0 л слюны. Ее pH колеблется от 5,25 до 8,0, а скорость секреции слюны у человека при «спокойном» состоянии слюнных желез составляет 0,24 мл/мин. Однако скорость секреции может колебаться даже в состоянии покоя от 0,01 до 18,0 мл/мин, что обусловлено раздражением рецепторов слизистой оболочки ротовой полости и возбуждением слюноотделительного центра под влиянием условных раздражителей. Слюноотделение при жевании пищи возрастает до 200 мл/мин. Количество и состав секрета слюнных желез меняется в зависимости от характера раздражителя. Слюна человека представляет собой вязкую, опалесцирующую, слегка мутную (благодаря присутствию клеточных элементов) жидкость с удельным весом 1,001—1,017 и вязкостью 1,10—1,33. Секрет смешанных всех слюнных желез человека содержит 99,4—99,5 % воды и 0,5—0,6 % плотного остатка, который состоит из неорганических и органических веществ (табл. 11.2). Неорганические компоненты в слюне представлены ионами калия, натрия, кальция, магния, железа, меди, хло- Таблица 11.2. Состав смешанного секрета всех слюнных желез человека
ра, фтора, йода, роданистых соединений, фосфата, сульфата, бикарбоната и составляют примерно 73 часть плотного остатка, а 2/3 приходится на органические вещества. Минеральные вещества слюны поддерживают оптимальные условия среды, в которой осуществляется гидролиз пищевых веществ ферментами слюны (осмотическое давление, близкое к нормальному, необходимый уровень pH). Значительная часть минеральных компонентов слюны всасывается в кровь слизистой оболочки желудка и кишечника (см. раздел 11.2.3). Это говорит об участии слюнных желез в поддержании постоянства внутренней среды организма. Органические вещества плотного остатка — это белки (альбумины, глобулины, свободные аминокислоты), азотсодержащие соединения небелковой природы (мочевина, аммиак, креатин), лизоцим и ферменты (альфа- амилаза и мальтаза). Альфа-амилаза является гидролитическим ферментом и расщепляет 1,4-глюкозидные связи в молекулах крахмала и гликогена с образованием декстринов, а затем мальтозы и сахарозы. Мальтаза (глюкозидаза) расщепляет мальтозу и сахарозу до моносахаридов. Вязкость и ослизняющие свойства слюны обусловлены наличием в ней мукополисахаридов (муцина). Слизь слюны склеивает частички пищи в пищевой комок; обволакивает слизистую оболочку ротовой полости и пищевода, она защищает ее от микротравм и проникновения патогенных микробов. Другие органические компоненты слюны, например холестерин, мочевая кислота, мочевина, являются экскретами, подлежащими удалению из организма. Слюна образуется как в ацинусах, так и в протоках слюнных желез. В цитоплазме железистых клеток содержатся секреторные гранулы, располагающиеся преимущественно в околоядерной и апикальной частях клеток, вблизи аппарата Гольджи. В ходе секреции размер, количество и расположение гранул изменяются. По мере созревания секреторных гранул они смещаются от аппарата Гольджи к вершине клетки. В гранулах осуществляется синтез органических веществ, которые двигаются с водой через клетку по эндоплазматической сети. В ходе секреции слюны количество коллоидного материала, находящегося в виде секреторных гранул, постепенно уменьшается по мере его расходования и возобновляется в период покоя в процессе его синтеза. В ацинусах слюнных желез осуществляется первый этап образования слюны. В первичном секрете содержится альфа-амилаза и муцин, которые синтезируются гландулоцитами. Содержание ионов в первичном секрете незначительно отличается от их концентрации во внеклеточных жидкостях, что говорит о переходе этих компонентов секрета из плазмы крови. В слюнных протоках состав слюны существенно изменяется по сравнению с первичным секретом: ионы натрия активно реабсорбируются, а ионы калия активно секретируются, но с меньшей скоростью, чем всасываются ионы натрия. В результате концентрация натрия в слюне снижается, тогда как концентрация ионов калия возрастает. Существенное преобладание реабсорбции ионов натрия над секрецией ионов калия увеличивает элек- тронегативность мембран клеток слюнных протоков (до 70 мВ), что вызывает пассивную реабсорбцию ионов хлора. Одновременно усиливается секреция ионов бикарбоната эпителием протоков, что обеспечивает ощелачивание слюны. Отделение слюны является сложным рефлекторным актом, вызванным раздражением рецепторов ротовой полости пищей или другими веществами {безусловно-рефлекторными раздражителями), а также раздражением зрительных и обонятельных рецепторов внешним видом и запахом пищи, видом обстановки, в которой происходит прием пищи (условно-рефлекторными раздражителями). Возбуждение, возникающее при раздражении механо-, хемо- и терморецепторов ротовой полости, достигает центра слюноотделения в продолговатом мозге по афферентным волокнам V, VII, IX, X пар черепно-мозговых нервов. Эфферентные влияния к слюнным железам поступают по парасимпатическим и симпатическим нервным волокнам. Преганлионарные парасимпатические волокна к подъязычным и подчелюстным слюнным железам находятся в составе барабанной струны (ветвь VII пары) к подъязычному и подчелюстному ганглиям, расположенным в теле соответствующих желез, а постганглионарные — от указанных ганглиев к секреторным клеткам и сосудам желез. К околоушным железам преганглионарные парасимпатические волокна идут от нижнего слюноотделительного ядра продолговатого мозга в составе IX пары черепно-мозговых нервов к ушному узлу, от которого постганглионарные волокна направляются к секреторным клеткам и сосудам. Прегангионарные симпатические волокна, иннервирующие слюнные железы, являются аксонами нейронов боковых рогов II—-VI грудных сегментов спинного мозга и заканчиваются в верхнем шейном ганглии. Отсюда постганглионарные симпатические волокна направляются к слюнным железам. Электрическое раздражение парасимпатических нервов в экспериментах на животных вызывает обильную секрецию жидкой слюны, содержащей небольшие количества органических веществ, а в гландулоци- тах уменьшается количество секреторных гранул. При стимуляции симпатических нервов из слюнных протоков выделяется небольшое количество густой и вязкой слюны с высоким содержанием ферментов и муцина, а в гландулоцитах увеличивается количество секреторных гранул. В связи с этим парасимпатические нервы называют секреторными, а симпатические — трофическими. При раздражении рецепторов слизистой оболочки рта пищевыми веществами у животных и человека парасимпатические влияния на слюнные железы преобладают над симпатическими, что вызывает обильную секрецию жидкой слюны. Во время приема пищи раздражаются тактильные, температурные и вкусовые рецепторы слизистой оболочки полости рта. Афферентные импульсы от них по чувствительным волокнам тройничного, языкоглоточного, лицевого и блуждающего нервов достигают слюноотделительного центра продолговатого мозга. При возбуждении верхнего слюноотделительного ядра центра эфферентные импульсы по преганглионарным парасимпатическим волокнам барабанной струны достигают подъязычного и нижнечелюстного ганглиев, где они переключаются на постганглионарные, парасимпатические волокна подъязычного нерва, иннервирующего подъязыч- ные и подчелюстные слюнные железы. При возбуждении нижнего слюноотделительного ядра центра эфферентные импульсы по преганглионарным парасимпатическим волокнам языкоглоточного нерва достигают ушного ганглия, где они переключаются на постганглионарные волокна ушновисочного нерва, иннервирующего околоушные слюнные железы. Под влиянием эфферентных импульсов в окончаниях парасимпатических постганглионарных волокон выделяется ацетилхолин, возбуждающий гландулоциты и расширяющий кровеносные сосуды железы. Поэтому секреторный эффект сопровождается повышением уровня кровоснабжения железы. Поступающие в продолговатый мозг афферентные импульсы возбуждают также чувствительные нейроны одиночного пучка, по аксонам которых сенсорные импульсы достигают ядер таламуса, где они переключаются на таламокортикальное пути и достигают коркового представительства вкусовой сенсорной системы (в области роландовой борозды). В коре больших полушарий сенсорная информация переключается на эфферентные корковые нейроны, аксоны которых передают эфферентные импульсы на парасимпатические и симпатические ядра гипоталамуса. Нисходящие влияния от парасимпатических ядер активируют бульбарный слюноотделительный центр, а от симпатических ядер — активируют преганглионарные симпатические нейроны II—V грудных сегментов спинного мозга, где возбуждение переключается на постганглионарные волокна, в окончаниях которых выделяется норадреналин. Центры слюноотделения рефлекторно могут не только возбуждаться, но и тормозиться. Например, при болевом раздражении, во время отрицательных эмоций и при умственном напряжении слюноотделение резко ослабляется или прекращается («пересыхает во рту»). Фармакологические вещества, относящиеся к группе холиномиметиков (например, пилокарпин, прозерин), усиливают слюноотделение, так как способствуют выделению ацетилхолина, а холинолитики — блокируют (например, атропин). Обильное отделение слюны наблюдается при асфиксии вследствие раздражения слюноотделительного центра угольной кислотой. 11.4.3. Жевание Жевание — физиологический акт, заключающийся в измельчении с помощью зубов пищевых веществ и формировании пищевого комка. Жевание обеспечивает качество механической обработки пищи и определяет время ее пребывания в полости рта, оказывает рефлекторное возбуждающее влияние на секреторную и моторную деятельность желудка и кишечника. В жевании участвуют верхняя и нижняя челюсти, жевательная и мимическая мускулатура лица, язык, мягкое небо. Механическая обработка пищи между верхними и нижними рядами зубов осуществляется благодаря перемещению нижней челюсти относительно верхней. У взрослого человека в ряду справа и слева имеются зубы разного функционального назначения — два резца и один клык (откусывающие пищу), два малых и три больших коренных, которые раздавливают и растирают пищу, — всего 32 зуба. Процесс жевания имеет четыре фазы — введения пищи в рот, ориентировочную, основную и формирования пищевого комка (рис. 11.10). Регуляция жевания осуществляется рефлекторно. Возбуждение от рецепторов слизистой оболочки рта (механо-, хемо- и терморецепторов) передается по афферентным волокнам II, III ветви тройничного, языкоглоточного, верхнего гортанного нерва и барабанной струны в центр жевания, ко- 1 1 11 А/1 111 1 л 6 IV | Л V 1 а в оо<| "Им» ко, И 'J L I J.I- ч— П11« I I» Г"* I <»—« »Я|< Рис. 11.10. Кимограмма жевательного периода. I — покой жевательной мускулатуры; II — фаза введения пищи в рот; III — ориентировочная фаза; IV — основная фаза; V — фаза формирования пищевого комка; а—б — опускание нижней челюсти, б—в — подъем нижней челюсти; о—о( — момент размалывания пищи. Под кимо- граммой — отметка времени — 1 с. торый находится в продолговатом мозге. Возбуждение от центра к жевательным мышцам передается по эфферентным волокнам тройничного, лицевого и подъязычного нервов. Возбуждение от чувствительных ядер ствола мозга по афферентному пути через специфические ядра таламуса переключается на корковый отдел вкусовой сенсорной системы, где осуществляется анализ и синтез информации, поступающей от рецепторов слизистой оболочки ротовой полости. На уровне коры больших полушарий происходит переключение сенсорных импульсов на эфферентные нейроны, которые по нисходящим путям посылают регулирующие влияния к центру жевания продолговатого мозга. 11.4.4. Глотание Глотание — рефлекторный акт, при помощи которого пища переводится из ротовой полости в желудок. Акт глотания состоит из трех фаз: ротовой (произвольной), глоточной (непроизвольной, быстрой) и пищеводной (непроизвольной, медленной). Пищевой комок (объемом 5—15 см3) скоординированными движениями мускулатуры щек и языка продвигается к его корню (за передние дужки глоточного кольца). Так завершается первая фаза глотания и начинается вторая. С этого момента акт глотания становится непроизвольным. Раздражение пищевым комком рецепторов слизистой оболочки мягкого неба и глотки передается по языкоглоточным нервам к центру глотания в продолговатом мозге. Эфферентные импульсы от него идут к мышцам полости рта, глотки, гортани и пищевода по волокнам подъязычных, тройничных, языкоглоточных и блуждающих нервов. Этот центр обеспечивает координированные сокращения мышц языка и мускулатуры, приподнимающей мягкое небо. Благодаря этому вход в полость носа со стороны глотки закрывается мягким небом, и язык перемещает пищевой комок в глотку. Одновременно происходит сокращение мышц, поднимающих нижнюю челюсть. Это приводит к смыканию зубов и прекращению жевания, а сокращение челюстно-подъязычной мышцы — к поднятию гортани. В результате осуществляется закрытие входа в гортань надгортанником. Этим предотвращается попадание пищи в дыхательные пути. В это же время открывается верхний пищеводный сфинктер, образованный волокнами циркулярного направления в верхней половине шейной части пищевода, и пищевой комок поступает в пищевод. Так начинается третья фаза. Верхний пищеводный сфинктер сокращается после перехода пищевого комка в пищевод, предотвращая пищеводно-глоточный рефлюкс (т. е. обратное поступление пищи в глотку). Затем осуществляется прохождение пищи по пищеводу и переход ее в желудок. Пищевод является мощной рефлексогенной зоной. Рецепторный аппарат представлен здесь в основном механорецепторами. Вследствие раздражения последних пищевым комком происходит рефлекторное сокращение мускулатуры пищевода. При этом последовательно сокращаются кольцевые мышцы (с одновременным расслаблением нижележащих). Волны перистальтических сокращений распространяются в сторону желудка, передвигая пищевой комок. Скорость их распространения 2—5 см/с. Сокращение мускулатуры пищевода связано с поступлением из продолговатого мозга эфферентной импульсации по волокнам возвратного и блуждающего нервов. Движение пищи по пищеводу обусловлено рядом факторов: во-первых, перепадом давления между полостью глотки и началом пищевода — от 45 мм рт. ст. в полости глотки (в начале глотания) до 30 мм рт. ст. (в пищеводе); во-вторых, наличием перистальтических сокращений мышц пищевода, в-третьих, тонусом мускулатуры пищевода, который в торакальном отделе почти в три раза ниже, чем в шейном, и, в-четвертых, силой тяжести пищевого комка. Скорость прохождения пищи по пищеводу зависит от консистенции пищи: плотная проходит за 3—9 с, жидкая —за 1 — 2 с. Центр глотания через ретикулярную формацию связан с другими центрами продолговатого и спинного мозга. Его возбуждение в момент глотания вызывает торможение деятельности дыхательного центра и снижение тонуса блуждающего нерва. Последнее вызывает задержку дыхания и учащение сердечных сокращений. Задержка дыхания предотвращает попадание пищи в дыхательные пути. При отсутствии глотательных сокращений вход из пищевода в желудок закрыт, так как мышцы кардиального отдела желудка находятся в состоянии тонического сокращения. Когда перистальтическая волна и комок пищи достигают конечной части пищевода, тонус мышц кардиальной части желудка рефлекторно снижается, и комок пищи поступает в желудок. При наполнении желудка пищей тонус мышц кардиального отдела желудка повышается и препятствует обратному поступлению желудочного содержимого из желудка в пищевод (желудочно-пищеводный рефлюкс). Пищеварение в желудке В желудке пища, смешанная со слюной и слизью, задерживается от 3 до 10 ч для ее механической и химической обработки. Желудок осуществляет следующие функции: 1) депонирование пищи\ 2) секрецию желудочного сока, обеспечивающего химическую обработку пищи; 3) перемешивание пищи с пищеварительными соками; 4) ее эвакуацию — передвижение порциями в двенадцатиперстную кишку; 5) всасывание в кровь небольшого количества веществ, поступивших с пищей; 6) выделение (экскрецию) вместе с желудочным соком в полость желудка метаболитов (мочевины, мочевой кислоты, креатина, креатинина), веществ, поступивших в организм извне (солей тяжелых металлов, йода, фармакологических препаратов); 7) образование активных веществ (инкрецию), принимающих участие в регуляции деятельности желудочных и других пищеварительных желез (гастрина, гистамина, соматостатина, мотилина и др.); 8) бактерицидное и бактериостатическое действие желудочного сока; 9) удаление недоброкачественной пищи, предупреждающее ее попадание в кишечник. 11.5.1. Секреторная функция желудка Секреторная функция желудка осуществляется желудочными железами, продуцирующими желудочный сок. Они состоят из трех видов клеток: главных, принимающих участие в выработке ферментов; обкладочных (париетальных), участвующих в выработке хлористоводородной (соляной) кислоты, и добавочных, выделяющих мукоидный секрет (слизь). В его состав входит также внутренний фактор кастла (гастромукопротен), участвующий в регуляции кровотворения. Натощак слизь выделяется также цилиндрическим эпителием, которым покрыта слизистая оболочка желудка. Железы кардиального отдела желудка секретируют в основном слизь. В железах пилорического отдела отсутствуют обкладочные клетки. Поэтому в секрете желез этого отдела отсутствует соляная кислота и его pH равен 7,8—8,4. Основную роль в желудочном пищеварении играют железы фундального отдела, включающего три секректорные зоны: дна, малой кривизны и тела желудка (рис. 11.11). Эти железы имеют все три типа клеток и выделяют основное количество желудочного сока. Состав желудочного сока. В состоянии покоя (натощак) из желудка человека можно извлечь около 50 мл желудочного содержимого нейтральной или слабокислой реакции (pH 6,0). Это смесь слюны и желудочного сока. Общее количество желудочного сока, отделяющегося у человека при обычном пищевом режиме, составляет 2,0—2,5 л в сутки. Это бесцветная,
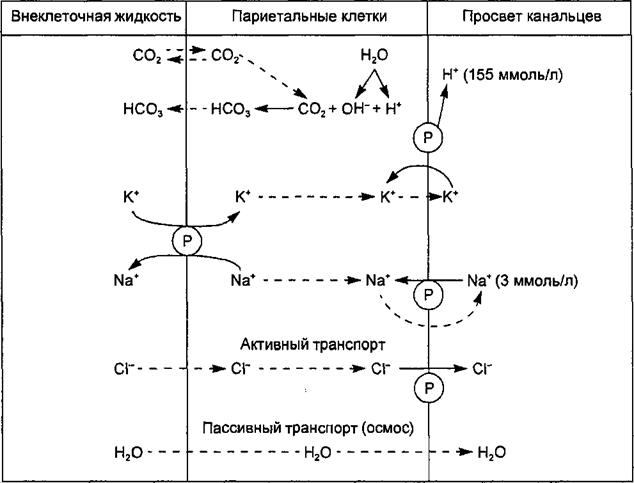
Рис. 11.11. Образование соляной кислоты желудочного сока. Пояснения в тексте. Символ ® означает активность ферментных транспортных систем мембраны кислотопродуцирующих клеток. Стрелками показано направление движения ионов и воды. прозрачная, слегка опалесцирующая жидкость с удельным весом 1,002— 1,007. В соке могут быть хлопья слизи. Желудочный сок имеет кислую реакцию (pH 0,8—1,5) вследствие высокого содержания в нем хлористоводородной (соляной) кислоты (0,3—0,5 %). Содержание воды в соке — 99,0— 99,5 %, а плотных веществ — 1,0—0,5 %. Плотный остаток представлен органическими и неорганическими веществами: хлоридами (5—6 г/л), сульфатами (10 мг/л), фосфатами (10—60 мг/л), гидрокарбонатами (0—1,2 г/л) натрия, калия, кальция и магния, аммиалом (20—80 мг/л). Значительная часть минеральных веществ всасывается в желудке и кишечнике в кровь и участвует в поддержании постоянства внутренней среды. Основной неорганический компонент желудочного сока — соляная кислота (см. ниже). Органическая часть плотного остатка состоит из ферментов и мукоидов (см. ниже). В небольшом количестве находятся в остатке азотсодержащие вещества небелковой природы (мочевина, мочевая кислота, молочная кислота и др.), подлежащие удалению из организма. Механизм секреции соляной кислоты. Хлористоводородная кислота вырабатывается париетальными (обкладочными) клетками желез желудка. Эти клетки характеризуются богатством митохондрий, расположенных вдоль внутриклеточных канальцев. Площадь мембраны канальцев и апикальной поверхности клеток во время стимуляции на высоте секреции резко возрастает за счет встроенных в мембрану тубовезикул (трубочек-пузырьков), что сопровождается значительным увеличением клеточных канальцев, проникающих вплоть до базальной мембраны. Это значительно увеличивает возможности синтеза гландулоцитом соляной кислоты. Вдоль канальцев располагается множество митохондрий, площадь внутренней мембраны которых возрастает в процессе биосинтеза НС1. Соответственно увеличивается площадь контакта канальцев и апикальной мембраны клетки. Таким образом, увеличение секреторной активности париетальных клеток обусловлено увеличением площади секреторной мембраны. Секреция НО является ярко выраженным цАМФ-зависимым процессом, активация которого протекает на фоне усиления гликогенолитической и гликолитической активности, что сопровождается продукцией пирувата. Окислительное декарбоксилирование пирувата до ацетил-КоА. СО2 осуществляется пируватдегидрогеназным комплексом и сопровождается накоплением в цитоплазме НАД • Н2. Последний используется для генерирования Н+ в процессе секреции НС1. Расщепление триглицеридов в слизистой оболочке желудка под влиянием триглицеридлипазы и последующая утилизация жирных кислот создает в 3—4 раза больший приток восстановительных эквивалентов в митохондриальную цепь переноса электронов. Как аэробный гликолиз, так и окисление жирных кислот запускаются посредством цАМФ-зависимого фосфорилирования соответствующих ферментов, обеспечивающих генерирование ацетил-КоА в цикле Кребса и восстановительных эквивалентов для электронпереносящей цепи митохондрий. Са2+ является необходимым элементом секреторной системы НС1. Процесс цАМФ-зависимого фосфорилирования обеспечивает активацию желудочной карбоангидразы, которая является регулятором кислотноосновного равновесия в кислотопродуцирующих клетках. Работа этих клеток сопровождается длительной и массовой потерей ионов Н+, что приводит к накоплению в клетке ОН-, способных оказать повреждающее действие на клеточные структуры. Нейтрализация гидроксильных ионов и является главной функцией карбоангидразы. Образующиеся бикарбонатные ионы посредством электронейтрального механизма выводятся в кровь, а ионы СГ входят в клетку. Кислотопродуцирующие клетки на наружных мембранах имеют две мембранные ферментные системы, участвующие в механизмах продукции Н+ и секреции НС1. Ими являются Ка+-К+-АТФаза и Н+-К+-АТФаза. Иа+-К+-АТФаза, расположенная в базолатеральных мембранах клеток, переносит К+ из крови в обмен на Na+, а Н+-К+-АТФаза, локализованная в секреторной мембране, транспортирует калий из первичного секрета в обмен на выводимые в желудочный сок ионы Н+. Процесс образования соляной кислоты кислотопродуцирующими клетками схематически представлен на рис. 11.11. В период секреции митохондрии всей массой охватывают в виде муфты секреторные канальцы, и их мембраны сливаются, образуя митохондриально-секреторный комплекс, где ионы Н+ непосредственно акцептируются Н+-К+-АТФазой секреторной мембраны и транспортируются из клетки. Таким образом, кислотообразующая функция обкладочных клеток осуществляется благодаря процессу фосфорилирования — дефосфорилирования, наличию митохондриальной окислительной цепи, транспортирующей ионы Н+ из матриксного пространства, а также активности Н+-К+-АТФазы секреторной мембраны, перекачивающей протоны из клетки за счет энергии АТФ. Вода поступает в канальцы клетки путем осмоса. Конечный секрет, поступающий в канальцы, содержит НС1 в концентрации 155 ммоль/л, хлористый калий в концентрации 15 ммоль/л и очень малое количество хлористого натрия. Роль соляной кислоты в пищеварении. В полости желудка хлористоводородная кислота: 1) стимулирует секреторную активность желез желудка; 2) способствует превращению пепсиногена в пепсин путем отщепления ингибирующего белкового комплекса; 3) создает оптимальную кислотность для действия протеолитических ферментов желудочного сока; 4) вызывает денатурацию и набухание белков (что способствует их расщеплению ферментами); 5) обеспечивает антибактериальный эффект секрета; 6) участвует в осуществлении механизма перехода пищи из желудка в двенадцатиперстную кишку, раздражая хеморецепторы ее слизистой оболочки; 7) участвует в регуляции секреции желудочных и поджелудочных желез, стимулируя образование гастроинтестинальных гормонов (гастрина, секретина); 8) возбуждает секрецию фермента энтерокиназы энтероцитами слизистой оболочки двенадцатиперстной кишки; 9) участвует в створаживании молока; 10) стимулирует моторную активность желудка. Ферменты желудочного сока и их роль в пищеварении. В полости желудка под влиянием протеолитических ферментов осуществляется начальный гидролиз белков до альбумоз и пептонов. Протеолитические ферменты желудочного сока обладают активностью в широком диапазоне колебаний pH с оптимумом действия при pH 1,5—2,0 и 3,2—4,0. Это обеспечивает гидролиз белков в условиях значительных колебаний концентрации соляной кислоты в желудочном соке, в слоях пищи, прилежащих к слизистой оболочке желудка, и в глубине содержимого желудка. В желудочном соке представлены семь видов пепсиногенов, объединенных общим названием пепсины. Образование пепсинов осуществляется из неактивных предшественников — пепсиногенов, находящихся в клетках желудочных желез в виде гранул зимогена. В просвете желудка пепсиноген активируется НС1 путем отщепления от него ингибирующего белкового комплекса. В дальнейшем в ходе секреции желудочного сока активация пепсиногена осуществляется аутокаталитически под действием уже образовавшегося пепсина. При оптимальной величине pH среды пепсин осуществляет гидролиз белков, разрывая в белковой молекуле пептидные связи, образованные группами фениламина, тирозина, триптофана и других аминокислот. В результате этого белковая молекула распадается на пептоны и пептиды. Пепсин обеспечивает гидролиз основных белковых веществ, особенно коллагена — основного компонента волокон соединительной ткани, К основным пепсинам желудочного сока относятся следующие. Пепсин А — группа ферментов, гидролизирующих белки при оптимуме pH 1,5—2,0. Часть пепсиногена (около 1 %) переходит в кровеносное русло, откуда вследствие небольшого размера молекулы фермента проходит через клубочковый фильтр в почках и выделяется с мочой (уропепсиноген). Определение содержания уропепсина в моче используется в лабораторной практике для характеристики протеолитической активности желудочного сока. Гастриксин (пепсин С), гидролизирующий белки при оптимуме pH 3,2— 3,5. Пепсин В (парапепсин) расщепляет желатину и белки соединительной ткани. При pH 5,6 и выше протеолитическое действие фермента ослабляется. Реннин (пепсин Д, химозин) расщепляет казеин молока в присутствии ионов Са2+. Желудочный сок содержит ряд непротеолитических ферментов. Среди них — желудочная липаза, расщепляющая жиры, которые находятся в пище в эмульгированном состоянии (жиры молока), на глицерин и жирные кислоты при pH 5,9—7,9. У грудных детей желудочная липаза расщепляет до 59 % жира молока. В желудочном соке взрослых людей липазы мало. Поэтому основное количество жиров переваривается в тонком кишечнике. Клетками поверхностного эпителия слизистой оболочки желудка вырабатывается лизоцим (муромидаза). Лизоцим обусловливает бактерицидные свойства желудочного сока. Уреаза расщепляет мочевину в желудке при pH 8,0. Освобождающийся при этом аммиак нейтрализует соляную кислоту и предотвращает избыточную кислотность химуса, поступающего из желудка в двенадцатиперстную кишку. Желудочная слизь и ее значение. Органическим компонентом желудочного сока является слизь. Нерастворимая слизь (муцин) является продуктом секреторной активности добавочных клеток и клеток поверхностного эпителия. Муцин выделяется через апикальную мембрану мукоцита, образует слой слизи толщиной 0,5—1,5 мм, он обволакивает слизистую оболочку желудка и препятствует повреждающему воздействию соляной кислоты и пепсинов на клетки слизистой оболочки и раздражающих веществ, поступивших с пищей. Этими же клетками одновременно с муцином продуцируется и бикарбонат. Образующийся при взаимодействии муцина и бикарбоната мукозо-бикар- бонатный барьер предохраняет слизистую оболочку от аутолиза под воздействием соляной кислоты и пепсинов. Слой слизи является преградой для обратной диффизии ионов водорода из полости желудка; он нейтрализует соляную кислоту благодаря буферным свойствам из-за наличия гидрокарбонатов, а также адсорбирует ферменты. Под влиянием длительного воздействия желчных кислот (при забрасывании их из двенадцатиперстной кишки), салицилатов, масляной и пропионовой кислот, алкоголя происходит нарушение слизистого барьера. Это приводит к обратной диффизии ионов водорода из полости желудка, контакту слизистой оболочки с пепсинами и ее повреждению в результате аутолиза. Так формируются пептические язвы желудка. Возникновению язвенного процесса способствуют продукты жизнедеятельности микроорганизма Helicobacter pylori, которые усиливают секрецию соляной кислоты. 11.5.2. Регуляция секреции желудочного сока Железы желудка в состоянии относительного покоя (в условиях отсутствия процесса пищеварения) выделяют небольшое количество сока нейтральной или слабощелочной реакции (фоновая секреция). Под влиянием пищевого раздражения во время приема пищи железы желудка секретируют значительный объем желудочного сока, богатого протеолитическими ферментами. Эта реакция желез является рефлекторным ответом на раздражение пищей рецепторов слизистой оболочки ротовой полости, глотки и желудка (безусловный рефлекс) и воздействие комплекса раздражителей, влияющих на другие рецепторы, предшествующих и сопутствующих приему пищи (условный рефлекс). Афферентные импульсы с раздражаемых пищей рецепторов (тактильных, температурных и вкусовых) по афферентным волокнам V, VII, IX и X черепно-мозговых нервов передаются в бульбарный, таламический, гипоталамический и корковый отделы пищевого центра. Нисходящие влияния коркового представительства пищевого центра активируют парасимпатические и симпатические ядра гипоталамуса. При возбуждении парасимпатических ядер их эфферентные влияния активируют клетки бульбарного отдела пищевого центра, импульсы от которых по преганглионарным волокнам блуждающего нерва передаются на постганглионарные нейроны интрамуральных ганглиев желудка (клетки Догеля I типа). Выделяемый в окончаниях их аксонов медиатор ацетилхолин через М-холинорецепторы мембраны гландулоцитов стимулирует секреторную деятельность главных, обкладочных и мукоидных клеток желез желудка. Поэтому после перерезки блуждающих нервов у собак не выделяется желудочный сок при раздражении пищей рецепторов полости рта, на ее вид и запах. Ацетилхолин активирует также эндокринные У-клетки слизистой оболочки антрального отдела желудка, вырабатывающие гастрин, который через кровоток достигает обкладочных клеток желез и через специальные рецепторы их мембраны вызывает выработку соляной кислоты. Ацетилхолин и гастрин в слизистой оболочке фундального отдела желудка стимулируют активность эндокринных клеток, выделяющих гистамин, который, диффундируя по межклеточным пространствам, через Н2- рецепторы мембраны обкладочных клеток стимулирует выработку большого количества кислого желудочного сока, бедного ферментами и мукоидами. После перерезки блуждающих нервов (в экспериментах на животных) сокогонный эффект ацетилхолина, гастрина и гистамина резко ослабляется. Поступление пищи в желудок во время еды усиливает безусловно-рефлекторное отделение желудочного сока, что обусловлено раздражением механорецепторов при растяжении его стенок (особенно антрального отдела) и хеморецепторов пептидами и экстрактивными веществами пищи. При этом рефлекс осуществляется с участием бульбарного отдела пищевого центра, дополняется местной реакцией, реализуемой через холинергические нейроны интраорганной нервной системы. Увеличение концентрации соляной кислоты в желудочном соке приводит к угнетению выработки гастрина У-клетками и последующим уменьшением кислотности сока. Этот механизм саморегуляции предотвращает избыточное выделение соляной кислоты. Подавляют секрецию соляной кислоты соматостатин, ВИП и серотонин (см. табл. 11.1). При возбуждении симпатических ядер гипоталамуса его нисходящие влияния стимулируют преганглионарные симпатические нейроны боковых рогов торакальных сегментов (IV—X) спинного мозга. Их эфферентные влияния стимулируют постганглионарные нейроны симпатических ганглиев, аксоны которых в составе чревных нервов достигают <
|
||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2020-12-09; просмотров: 175; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.147 (0.021 с.) |