Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Физиологические гомеостатические механизмыСодержание книги
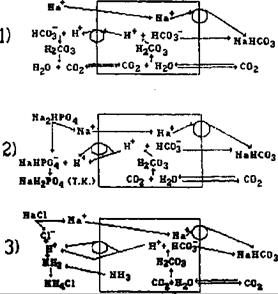
Поиск на нашем сайте Эти механизмы, включающие функции легких, почек, желудочно-кишечного тракта, печени и костной ткани, обеспечивают нейтрализацию, связывание и выведение из организма Н+ и ОН" ионов в составе различных соединений, в зависимости от особенностей метаболизма и состояния буферных систем внутренней среды. 15.3.1. Легкие и кислотно-основное состояние Процессы газообмена между кровью и окружающим воздухом, происходящие в легких, обеспечивают регуляцию кислотно-основного состояния, поскольку внешнее дыхание — это основной путь удаления из крови летучей угольной кислоты. При физиологических условиях в покое в организме вырабатывается 10 ммоль/мин угольной кислоты, а выделяется через легкие 13 000—15 000 ммолей СО2 в сутки. Благодаря бикарбонатному буферу образующиеся при метаболизме кислоты не вызывают закисления крови, а приводят к увеличению содержания СО2 в крови с последующим удалением через легкие. Удаленный из плазмы углекислый газ немедленно восполняется за счет двух механизмов, происходящих в крови легочных капилляров. Во-первых, образование оксигемоглобина ведет к распаду карбаминовой связи гемоглобина и освобождению СО2 из эритроцитов; во-вторых, под влиянием карбангидразы, фермента, катализирующего реакцию: Н2СО3 Н2О + СО2 Количество выводимого из организма углекислого газа зависит от глубины и частоты дыхания, т. е. от альвеолярной вентиляции. Величина же альвеолярной вентиляции регулируется дыхательным центром, деятельность которого зависит или даже определяется парциальным напряжением СО2 в крови и pH внеклеточной жидкости мозга. Таким образом, усиление дыхания и альвеолярной вентиляции происходит при увеличении напряжения в крови СО2 и уменьшении pH. Нарастающее при этом удаление углекислоты через легкие приводит ее содержание в крови в соответствие с концентрацией бикарбоната. Следовательно, легкие выступают в роли физиологической гомеостатической системы, не только выводящей избыток летучей кислоты, но и восстанавливающей емкость буферных систем, прежде всего бикарбонатной и гемоглобиновой. При быстром поступлении во внутреннюю среду оснований вентиляция легких уменьшается и количество выделяемого СО2 существенно снижается. Концентрация углекислоты возрастает и pH поддерживается на исходном уровне. 15.3.2. Почки и кислотно-основное состояние Почки осуществляют выведение из внутренней среды организма нелетучих органических и неорганических кислот. Избыток кислот при этом может выводиться как в свободном состоянии, так и в виде нейтральных солей. Сильные неорганические кислоты выделяются из внутренней среды только через почки и только в связанной форме. В физиологических условиях почки выделяют кислую мочу, pH которой колеблется от 5 до 7. Однако, в зависимости от кислотно-основного состояния внутренней среды, моча может быть более кислой (pH 4,0) или даже щелочной (pH 8,0). Участие почек в регуляции кислотно-основного состояния внутренней среды обеспечивается совокупностью взаимосвязанных процессов, происходящих в структурах нефрона. К их числу относятся: 1) секреция водородных ионов в мочу клетками канальцевого эпителия, 2) реабсорбция (обратное всасывание) профильтровавшихся в мочу оснований, прежде всего бикарбоната, пополняющих резерв в крови, 3) образование в клетках эпителия канальцев ионов бикарбоната, больше или меньше всасываемого в кровь, 4) образование и диффузия в мочу аммиака, создающего катион аммония, присоединяя Н-ион, 5) фильтрация в первичную мочу из плазмы крови соединений, обладающих кислыми или щелочными свойствами, 6) обмен ионов СГ, Na+, К+, фосфатов, сульфатов и двухвалентных катионов (Са2+, Mg2+). Важнейшую роль в способности почек к выделению кислой мочи играет секреция водородных ионов клетками эпителия проксимальных и дистальных канальцев (рис. 15.2). Эта способность обусловлена наличием в клетках эпителия фермента карбоангидразы, обеспечивающего образование из СО2 и Н2О угольной кислоты, с последующей диссоциацией: н2о + со2 -> н2со3 н+ + нсо;. Ион водорода активно секретируется с затратой энергии через апикальную мембрану клетки эпителия в просвет канальца, а из канальцевой мочи в клетку диффундирует ион натрия, обеспечивая равновесие электрических зарядов. Из клетки, уже через базальную мембрану, Na+ активно с помощью Na-K-насоса удаляется в интерстиций и кровь, туда же пассивно по градиенту концентрации поступает НСО7 образуя бикарбонат натрия. Следовательно, секреция Н-ионов обеспечивает обратное всасывание бикарбоната и натрия, причем основная часть секретируемых ионов водорода тратится именно на обеспечение реабсорбции бикарбоната. Так, секретируемый Н-ион в просвете проксимального канальца взаимодействует с профильтровавшимся в мочу из плазмы ионом бикарбоната (в сутки около 5000 ммоль), образуя угольную кислоту. Под влиянием фермента карбоангидразы, встроенного в мембрану щеточной каемки клеток эпителия канальца, угольная кислота разлагается на СО2 и воду: карбоангидраза Н+ 4- НСО7 -> Н2СО3 > СО2 + Н2О. Молекулы углекислого газа легко диффундируют в клетки, где реакция катализируется в обратном направлении. Таким образом, на суточ-
Кружки обозначают мембранные транспортные системы. Первый фрагмент (сверху) — секреция Н+-ионов в мочу клетками эпителия проксимальных канальцев обеспечивает реабсорбцию профильтровавшегося в первичную мочу натрия и транспорт во внутреннюю среду бикарбоната натрия, что способствует сохранению нормального значения величины актуального бикарбоната крови. Второй (средний) фрагмент — секреция водородных ионов в мочу ведет к реабсорбции натрия из диссоциирующих солей неорганических кислот и появлению титруемых кислот мочи. Третий фрагмент — аммониогенез как путь выведения избытка Н+-ионов. Секретируемые эпителием в просвет канальца аммиак и Н-ионы взаимодействуют между собой и с ионом хлора, образуя экскретируемый хлорид аммония. ную реабсорбцию до 5000 ммоль бикарбоната расходуется эквивалентное количество секретированных ионов водорода. Выделяется же с мочой только 60 ммоль/сут ионов водорода, преимущественно в связанном виде. Наибольшая часть секретированных канальцами Н+ (65—75 %) участвуют в процессе аммониогенеза, обеспечивающего выведение из плазмы крови анионов сильных неорганических и органических кислот в виде аммонийных солей. Клетки проксимальных и дистальных почечных канальцев, благодаря процессам дезаминирования и дезамидирования аминокислот, образуют аммиак. Основным источником является глутамин, который дезаминируется под влиянием фермента глутаминазы, 40 % аммиака образуется из аминокислот аланина и глицина. Аммиак, благодаря высокой растворимости в липидах, легко диффундирует в просвет канальца, где присоединяет секретированные Н+ и образует ион аммония; NH3 + Н+ -» NH^. Ионы аммония вытесняют натрий из соединений с анионами сильных кислот, натрий реабсорбируется, а анионы кислот в виде аммонийных солей выделяются с мочой. Содержащиеся в крови кислые и щелочные компоненты буферных систем, например одно- и двухосновный фосфаты, бикарбонат, а также слабые органические кислоты — молочная, лимонная, р-оксимаслянная и др., так же как и анионы сильных неорганических кислот (СГ, SO^), фильтруются из плазмы крови в первичную мочу. Часть из них реабсорбируется в проксимальных канальцах, особенно существенно бикарбонат (до 80— 90 %), мочевая кислота (более 90 %). Но наряду с обратным всасыванием многие из указанных веществ, особенно органические кислоты и основания (холин), активно секретируются клетками канальцевого эпителия. Таким образом, уровень этих веществ в моче определяется соотношением трех основных процессов: клубочковой фильтрации, канальцевых реабсорбции и секреции. Секретированные в мочу водородные ионы участвуют в образовании титруемых кислот мочи. Основную роль при этом играют фосфаты. Поскольку в плазме крови в 4 раза больше двухосновных фосфатов, поэтому и в первичную мочу их фильтруется соответственно больше. Под воздействием секретированного эпителием канальца Н-иона происходит перевод двухосновного фосфата в одноосновный, путем ионного обмена протона на натрий: Na2HPO4 + Н+ -> NaH2PO4 + Na+ Освобождаемый ион Na+ реабсорбируется в канальцах в кровь. Таким образом, этот процесс способствует сохранению во внутренней среде натрия и удалению из нее избытка Н+-ионов. Многие органические кислоты содержатся в плазме крови в виде солей и, фильтруясь в клубочках, поступают в первичную мочу. Под воздействием секретированных канальцами Н-ионов в моче происходит образование свободных слабых, т. е. плохо диссоциирующих, кислот, также составляющих титруемую кислотность мочи. Титруемой кислотностью мочи называют сумму Н-ионов слабых кислот и солей, определяемую путем титрования мочи щелочью до величины pH, равной pH крови. Эта величина показывает, насколько больше водородных ионов содержится в моче по сравнению с плазмой крови, т. е. насколько эффективно почки поддерживают постоянство pH. В клетках эпителия дистальных отделов нефрона кроме описанной выше секреции водородных ионов происходит секреция ионов К+, при этом ионы Н+ и К+ конкурируют в обмене на Na+. В связи с этим выведение калия и протона может изменяться противоположно друг другу. Так, при избытке водородных ионов секреция калия снижается, а при недостатке — возрастает. Напротив, избыток К+ снижает секрецию водородных ионов, а недостаток К+ повышает ее. Недостаток калия стимулирует и синтез аммиака в эпителиальных клетках канальцев для выведения избытка Н-ио- нов в виде аммонийных солей. Однако секреция протонов и калия связана с транспортом натрия и меняется пропорционально сдвигам реабсорбции натрия. При снижении дистальной реабсорбции натрия соответственно снижается секреция Н+ и К+ ионов, напротив, усиление всасывания натрия в дистальных канальцах приводит к повышению секреции калия и водородных ионов. При поступлении в кровь избытка кислот или оснований компенсация сдвига кислотно-основного состояния осуществляется прежде всего буферными системами крови, дыхательная компенсация формируется через 16— 18 ч, а почечная компенсация развивается еще медленнее — спустя 2— 3 сут. При этом почечные механизмы обладают значительной инерционностью и даже после удаления из организма избытка кислот или оснований возвращение к исходному кислотно-основному состоянию продолжается также около 2—3 сут. 15.3.3. Желудочно-кишечный тракт, печень, костная ткань и кислотно-основное состояние Характер питания человека и особенности пищеварения в значительной мере сказываются на кислотно-основном состоянии его внутренней среды, В пищевых продуктах растительного происхождения содержится большое количество солей органических кислот (щавелевой, яблочной, лимонной, янтарной и др.), после окисления которых и выведения кислотных валентностей с углекислотой через легкие остаются и накапливаются связанные с катионами щелочные валентности. Поэтому растительная пища способствует ощелачиванию внутренней среды, соответственно, основания в большем количестве выводятся с мочой, что увеличивает pH мочи и она может приобретать щелочную реакцию. Белковая пища животного происхождения способствует закислению внутренней среды, так как в составе животных белков содержатся сера и фосфор, что приводит к образованию сильных неорганических кислот. Повышенное выведение их с мочой приводит к снижению pH мочи и она приобретает резко кислую реакцию. Всасывание в желудочно-кишечном тракте кислот или щелочей, содержащихся в различных напитках, в том числе алкогольных и минеральных водах, также приводит к изменению кислотно-основного состояния внутренней среды. Сравнительно небольшую роль в регуляции кислотно-основного состояния играют пищеварительные функции желудка и кишечника. Париетальные клетки слизистой оболочки желудка секретируют соляную кислоту. Водородные ионы образуются в клетках под влиянием карбоангидразы, анион хлора поступает в клетки из хлористого натрия крови путем обмена на бикарбонат. Однако при этом ощелачивания крови не происходит, так как СГ желудочного сока в кишечнике всасывается обратно в кровь. Значительное количество бикарбонатов поступает в кишечник в составе сока поджелудочной железы и кишечного сока. При этом ионы Н+ всасываются в кровь. Обратное всасывание претерпевают и бикарбонатные ионы, поэтому сдвигов кислотно-основного состояния крови не происходит. При кислой диете увеличивается выделение двухвалентных катионов Са2+ и Mg2+ а при щелочной — и одновалентных катионов. Хотя значение желудочно-кишечного тракта в физиологической регуляции кислотно-основного состояния невелико, тем не менее при изменениях, а тем более нарушениях функций желудка и кишечника неизбежно происходят сдвиги и кислотно-основного состояния. Стойкое и резкое повышение кислотности желудочного сока может вести к увеличение щелочного резерва крови из- за потерь Н-ионов и избыточного всасывания бикарбоната. Частая рвота, особенно при непроходимости кишечника, вызывая потерю Н+-ионов и хлоридов, вызывает гипохлоремию и защелачивание внутренней среды, что требует обязательной врачебной коррекции. В печени происходит утилизация молочной кислоты с образованием гликогена, здесь разрушаются кетоновые тела (ацетоуксусная и бета-окси- масляная кислоты), происходят метаболические превращения органических кислот и оснований. Образуя желчь с изменяемой степенью щелочной реакции, печень способствует удалению из внутренней среды разных количеств оснований. В нейтрализации кислот внутренней среды принимает участие костная ткань, поскольку в ней связано огромное количество катионов и фосфата. При длительном накоплении в крови кислот и снижении pH внеклеточной жидкости происходит остеолиз и вымывание из скелета фосфата кальция. В результате реакции с угольной кислотой образуются бикарбонатный и однозамещенный фосфатный анионы, нейтрализующие избыток Н-ионов. Благодаря огромным запасам в скелете фосфата кальция этот механизм обладает высокой эффективностью, выход из скелета 1 моль фосфата кальция нейтрализует и способствует в дальнейшем выведению из организма через почки 4 эквивалентов кислоты. Однако потеря костной тканью фосфата кальция вызывает деминерализацию скелета и потерю его прочности. В костной ткани находится до 45 % всего натрия, содержащегося в организме, однако выход натрия из костной ткани возможен только при резорбции кости, например, в условиях кислой реакции среды. В таких случаях натрий костей, поступая во внеклеточную среду, способствует нейтрализации кислотного сдвига.
|
||||||
|
|
Последнее изменение этой страницы: 2020-12-09; просмотров: 132; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.220 (0.014 с.) |