Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Функции промежуточного мозгаСодержание книги
Поиск на нашем сайте
К промежуточному мозгу относят две структуры: зрительные бугры (таламус), расположенные с обеих сторон III желудочка мозга, и гипоталамус (подбугорье), находящийся книзу от III желудочка. Функция таламуса состоит в распределении потоков афферентной информации, следующей к коре больших полушарий. Таламус участвует в восприятии сенсорных сигналов и формировании целенаправленного поведения. Гипоталамус является высшим центром регуляции вегетативных функций. 4.7.1. Функции таламуса Таламус расположен между большими полушариями и мозговым стволом, в нем происходят переключение и переработка афферентных сигналов, поступающих к коре. В таламусе насчитывают до 120 ядер, среди которых различают специфические, или релейные, ядра (подразделяются на сенсорные и моторные), неспецифические ядра, а также ассоциативные ядра. Специфические ядра таламуса содержат две основные группы клеток. Во-первых, релейные нейроны с ветвящимися в пределах ядра дендритами и длинным аксоном, образующим моносинаптическую проекцию на нейроны IV и III слоев коры. Во-вторых, локальные возбуждающие и тормозные интернейроны, аксоны которых не выходят за пределы ядра и участвуют в образовании локальной сети. Ассоциативные ядра имеют такую же нейронную организацию, а в неспецифических ядрах основная масса нейронов похожа на клетки ретикулярной формации ствола. Основной характеристикой всех релейных ядер является специализация афферентных входов от периферических рецепторов и чувствительных ядер ствола и высокоточная передача нервных импульсов к проекционной области коры с минимальным искажением входного сигнала. Так, например, наружные коленчатые тела получают информацию от сетчатки и передают ее в первичную зрительную кору (затылочные доли). Медиальные коленчатые тела относятся к слуховой сенсорной системе и образуют проекции на первичную слуховую кору (височные доли), а вентробазалъные ядра, получающие афферентную информацию от поверхности тела и от мышц,— на соматосенсорную кору (постцентральные извилины). При переключении сигналов в сенсорных ядрах сохраняется топографическая разграниченность проекций. Каждому рецептивному полю сетчатки соответствуют определенные переключательные нейроны латерального коленчатого тела (ретинотопическая организация). Так, например, нейроны вентральной части латерального коленчатого тела получают афферентную информацию от наружных участков сетчатки, а дорсальные нейроны — от внутренней области сетчатки. После этого таламические нейроны в том же порядке передают информацию в проекционную зрительную кору, в которой пространственное распределение возбужденных нейронов повторяет топографическую организацию сетчатки и действующих на нее зрительных стимулов. Это соответствие создает в зрительной коре нейронную карту зрительного поля.
Рецептивные поля разных участков тела соматотопически представлены на нейронах вентробазального ядра. Каждый участок поверхности лица, туловища, конечностей имеет сенсорный путь к определенным нейронам этого ядра. По тому же принципу осуществляется проекция от разных участков кортиева органа внутреннего уха на определенные нейроны внутреннего коленчатого тела. Такая организация называется тонотопической, она позволяет передавать независимо друг от друга сигналы от слуховых рецепторов, различающихся по своей чувствительности к звукам разной высоты. Передача информации о разных качествах одного стимула происходит параллельно и осуществляется разными нейронами сенсорного ядра. Так, например, магноцеллюлярный путь от больших рецептивных полей периферии сетчатки используется для передачи информации о положении объектов в зрительном поле и об их движении, а парвоцеллюлярный путь от малых рецептивных полей центральной ямки сетчатки предназначен для передачи информации о форме и цвете объектов. В наружном коленчатом теле переключение сигналов каждого параллельного пути происходит независимо. По такому же принципу, т. е. параллельно, с участием разных нейронов вентробазального ядра передается соматосенсорная чувствительность от тактильных рецепторов кожи и проприоцепторов мышц, сухожилий и суставов. Нейроны таламуса не объединяют эту информацию, переправляя ее в разные регионы проекционной области, которой является соматосенсорная кора. Моторные ядра таламуса, среди которых самым крупным является вентролатеральное ядро, обеспечивают переключение афферентации от базальных ганглиев и мозжечка к коре, что необходимо для создания двигательных программ. В оральном отделе вентролатерального ядра переключаются импульсы, следующие от внутренних областей бледного шара к вторичной моторной коре, а в каудальном отделе проецируются афференты от ядер мозжечка и красного ядра, которые переключаются на релейные нейроны, связанные с первичной моторной корой. В медиокаудальной части вентролатерального ядра переключаются сигналы, следующие от вестибулярных ядер и мышечных веретен во вторичную моторную кору, а в ростральном отделе афференты от наружных областей бледного шара проецируются на нейроны, имеющие выход в первичную и во вторичную моторную кору.
Ассоциативные ядра отличаются от специфических релейных ядер тем, что не относятся к какой-либо одной сенсорной системе, а большую часть своих афферентов получают от других ядер таламуса, как релейных, так и неспецифических. Функциональная связь проекционных и ассоциативных ядер способствует интеграции всех этапов переработки сенсорной информации, необходимой для комплексного восприятия сенсорных стимулов. Интегративная функция ассоциативных ядер состоит в объединении деятельности всех ядер таламуса с ассоциативными областями коры. Каждое ассоциативное ядро имеет проекцию на определенную ассоциативную область коры: медиодорсалъное ядро образует проекцию на орбитофронтальную кору, группа латеральных ядер — на теменную кору, а группа задних ядер—на височную. Упорядоченное распределение информационных потоков отражает функциональную специализацию указанных областей коры. Характерным признаком ассоциативных ядер является полимодальная (полисенсорная) конвергенция афферентных входов от двух и более сенсорных систем, а также от гипоталамуса и структур лимбической системы мозга. Среди нейронов ассоциативных ядер имеются клетки, которые возбуждаются только в ответ на действие определенного сочетания сенсорных стимулов. Полисенсорная конвергенция позволяет объединять в таламусе разные виды сенсорной информации для комплексного восприятия окружающего мира, а включение в этот процесс лимбических структур формирует субъективное эмоциональное отношение к полученной информации. Неспецифические ядра получают афферентные сигналы преимущественно из двух источников: ретикулярной формации и спиноталамического тракта. Помимо этого неспецифические ядра получают афферентные входы от гипоталамуса, лимбических структур, базальных ганглиев, специфических и ассоциативных ядер таламуса. В результате неспецифические ядра оказываются морфологически и функционально связанными со многими системами, что определяет их активность, зависящую не столько от определенной модальности, сколько от общей суммы афферентных сигналов. При такой организации точность проекции неспецифических ядер на кору не имеет значения, их проекции диффузно распространяются практически на все области коры, а также на лимбическую систему и гипоталамус. Влияние неспецифических ядер на кору состоит в модуляции активности ее нейронов, т. е. прямо не возбуждает и не тормозит, но изменяет их возбудимость при получении специфической афферентации. Этот механизм используется для регуляции функциональной активности мозга, которая изменяется на протяжении суток.
Таламус, благодаря организации своих связей с другими регионами мозга, вовлекается в осуществление различных функций: формирование эмоций (совместно с лимбической системой), чувственное восприятие (совместно с сенсорными областями коры), осуществление психической деятельности (совместно с ассоциативными регионами коры). 4.7.2. Функции гипоталамуса Гипоталамус является высшим центром регуляции вегетативных функций, он координирует деятельность симпатического и парасимпатического отделов вегетативной нервной системы и согласует ее с двигательной активностью человека. Одновременно гипоталамус управляет секрецией гормонов гипофиза, и тем самым контролирует эндокринную регуляцию внутренних процессов. Ядра гипоталамуса являются центрами регуляции водносолевого баланса, температурного гомеостазиса, чувства голода и насыщения, полового поведения, выполняют функцию координатора биологических ритмов. Будучи важнейшей мотивационной структурой мозга, гипоталамус участвует в формировании биологических мотиваций, эмоций и организации целенаправленного поведения. 4.7.2.1. Роль гипоталамуса в регуляции вегетативных функций Гипоталамус оперативно получает информацию от интерорецепторов внутренних органов, сигналы от которых поступают через ствол мозга, кроме того в гипоталамус поступает часть сенсорных сигналов от поверхности тела и некоторых специализированных сенсорных систем (зрительная, обонятельная, слуховая). Некоторые нейроны гипоталамуса имеют собственные осмо-, термо- и глюкорецепторы, которые воспринимают сдвиги осмотического давления, температуры и уровня глюкозы в ликворе и крови. В отличие от периферических рецепторов они называются центральными, а их наличие именно в гипоталамусе связано со слабой выраженностью гематоэнцефалического барьера. Гипоталамус имеет двусторонние связи с лимбической системой мозга, ретикулярной формацией и корой больших полушарий, что позволяет ему координировать вегетативные функции с определенным поведением, например с переживанием эмоций. Гипоталамус образует проекции на вегетативные центры ствола и спинного мозга, что позволяет ему осуществлять прямой контроль деятельности этих центров. Он регулирует ключевые механизмы эндокринной регуляции вегетативных функций, осуществляя управление секрецией гормонов гипофиза. Необходимые для вегетативной регуляции переключения осуществляют нейроны ядер гипоталамуса, в разных классификациях их насчитывают от 16 до 48. Искусственное раздражение гипоталамуса электрическим током через вживленные электроды у экспериментальных животных сопровождается разными комбинациями ответных вегетативных и поведенческих реакций в зависимости от того, какая область гипоталамуса подвергалась раздражению. Так, например, при стимуляции задней области гипоталамуса происходит повышение кровяного давления, увеличение частоты сокращений сердца, учащение и углубление дыхания, расширение зрачков. Одновременно с этим поднимается шерсть, выпускаются когти, изгибается горбом спина и оскаливаются зубы. Вегетативные сдвиги свидетельствуют об активации симпатического отдела, а поведение является аффективнооборонительным. Эта область гипоталамуса получила название эрготроп- ной, т. е. способствующей мобилизации энергии.
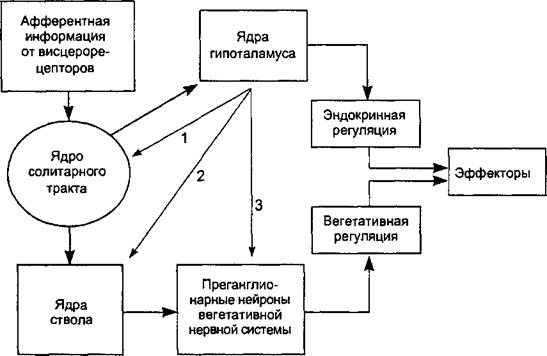
Раздражение ростральных отделов гипоталамуса и преоптической области вызывало у тех же животных пищевое поведение: они начинали поглощать пищу, даже если были досыта накормлены. У них увеличивалось выделение слюны, усиливалась моторика желудка и кишечника, а частота сердечных сокращений и дыхания уменьшалась, становился меньшим и мышечный кровоток, что характерно для повышения парасимпатического тонуса. Эта область гипоталамуса была названа трофотропной. Таким образом, активация разных областей гипоталамуса запускает запрограммированные комплексы поведенческих и вегетативных реакций. Задача гипоталамуса заключается в том, чтобы оценить поступающую к нему из разных источников информацию и на ее основе выбрать тот или иной комплекс ответных реакций, объединяющий поведение с определенной активностью обоих отделов вегетативной нервной системы. Следует обратить внимание, что не только уже произошедшие отклонения параметров гомеостазиса, но и любое потенциально угрожающее ему событие может активировать деятельность гипоталамуса (опережающее реагирование). Так, например, при внезапном возникновении опасной ситуации, угрожающей жизни, вегетативные сдвиги у человека (увеличение частоты сокращений сердца, повышение кровяного давления и т. п.) происходят быстрее, чем он обратится в бегство, т. е. такие сдвиги уже учитывают характер последующей мышечной активности. Непосредственный контроль тонуса вегетативных центров, а значит и выходной активности вегетативной нервной системы, гипоталамус осуществляет с помощью эфферентных связей с тремя важнейшими областями (рис. 4.27). Во-первых, он регулирует активность нейронов ядра солитарного тракта, которое является главным адресатом сенсорной информации от внутренних органов, участвует в контроле температуры, кровообращения, дыхания и взаимодействует с ядром блуждающего нерва и других парасимпатических нейронов. Во-вторых, гипоталамус определяет актив-
Рис. 4.27. Схема гипоталамического контроля вегетативных функций. 1. Эфферентный путь от гипоталамуса к ядру солитарного тракта.
2. Эфферентный путь от гипоталамуса к ростральной вентральной области продолговатого мозга. 3. Эфеерентный путь от гипоталамуса к вегетативным нейронам спинного мозга.
ность ростральной вентральной области продолговатого мозга, имеющей решающее значение в повышении общей выходной активности симпатического отдела. Эта активность проявляется в повышении кровяного давления, увеличении частоты сокращений сердца, секреции потовых желез, расширении зрачков и сокращении мышц, поднимающих волосы. В третьих, некоторые нейроны гипоталамуса образуют прямые проекции на вегетативные нейроны спинного мозга, что позволяет быстро изменять характер их активности. 4Л.2.2. Роль гипоталамуса в регуляции эндокринных функций Группы мелкоклеточных нейронов гипоталамуса способны действовать как нейроэндокринные преобразователи, трансформирующие кратковременные нервные импульсы в длительные гуморальные влияния. В области воронки мозга секретируемые мелкоклеточными нейронами гипоталамуса пептиды по системе портальных вен поступают в аденогипофиз (рис. 4.28) и регулируют образование его гормонов. Гипофизотропные нейроны гипоталамуса получают информацию от разветвленной афферентной сети волокон, передающих сигналы от лимбической системы, ствола, зрительной, слуховой и обонятельной сенсорных систем. В результате интеграции этой обширной информации формируется выходной сигнал в виде необходимой порции нейрогормонов. Две разновидности крупноклеточных нейронов гипоталамуса, находящиеся в супраоптическом и паравентрикулярном ядрах, синтезируют вазопрессин и окситоцин. Эти нейропептиды доставляются аксонным транспортом к окончаниям нейронов в задней доле гипофиза, где депонируются и
Рис. 4.28. Схема гипоталамического контроля деятельности гипофиза. 1. Нейроны супраоптического и паравентрикулярного ядер гипоталамуса образуют вазопрессин и окситоцин, которые выделяются в задней доле гипофиза в кровь. 2 и 3. Нейроны, секретирующие либерины и статины в кровеносные капилляры; через портальные вены эти пептиды поступают к эндокринным клеткам аденогипофиза. 4 и 5. Нейроны, регулирующие деятельность нейросекреторных клеток гипоталамуса.
откуда выделяются в кровь, чтобы действовать в качестве гормонов. Крупноклеточные нейроны гипоталамуса выполняют функцию нейроэндокринного преобразователя, в котором количество выделяемых нейрогормонов увеличивается пропорционально электрической активности таких клеток.
|
|||||||||
|
|
Последнее изменение этой страницы: 2020-12-09; просмотров: 151; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.15.4.70 (0.015 с.) |