Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Рефлекторная регуляция функций организмаСодержание книги
Поиск на нашем сайте
Рефлекс — стереотипная реакция организма в ответ на раздражитель, реализуемая с помощью нервной системы. Вызывающие рефлексы раздражители могут иметь как физическую (механические, электрические, температурные, звуковые, световые и т. п. раздражители), так и химическую природу. Структурной основой рефлекса является рефлекторная дуга, представляющая собой совокупность морфологически взаимосвязанных образований, обеспечивающих восприятие, передачу и переработку сигналов, необходимых для реализации рефлекса. Рефлекторная дуга по своему строению и назначению элементов представляет собой описанный выше контур регуляции. Она включает следующие звенья (рис. 3.4): 1) сенсорные рецепторы (датчики), воспринимающие стимулы внешней или внутренней среды, 2) афферентные, или чувствительные, нервные проводники (каналы сигналов входа), 3) нервные центры (аппарат управления), состоящие из афферентных, промежуточных, или вставочных, и эфферентных нейронов, т. е. по-

Раздражение рецептора ведет к поступлению нервного импульса по афферентному проводнику к афференнтному нейрону спинального ганглия, а от него к вставочному и двигательному нейронам спинного мозга (нервный центр рефлекса). От двигательного нейрона передних рогов спинного мозга по эфферентному проводнику нервный импульс передается к эффектору — скелетной мышце. Сокращение последней вызывает раздражение расположенных в мышце рецепторов и по афферентным проводникам информация о сокращении поступает обратно в нервный центр (канал обратной связи)
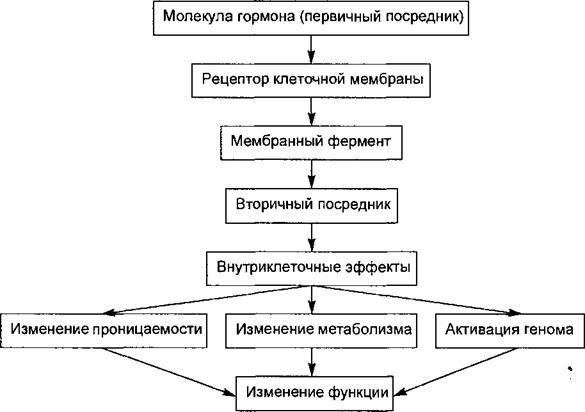
4) 5) эффекторы, или исполнительные органы (объекты управления). Однако для оптимальности регуляции необходима информация о реакциях эффектора на управляющие сигналы, в связи с чем обязательным звеном рефлекторного акта является канал обратной связи. Таким образом, структурную основу рефлекса лучше называть не рефлекторной дугой, а рефлекторным кольцом. Рецепторами называют специализированные образования, предназначенные для восприятия клетками или нервной системой различных по своей природе стимулов или раздражителей. Различают два типа рецепторов — сенсорные, т. е. обеспечивающие восприятие нервной системой различных раздражителей внешней или внутренней среды, и клеточные химические рецепторы — специальные структуры мембран, обеспечивающие восприятие информации, переносимой молекулами химических веществ —- медиаторов, гормонов, антигенов и т. п.
3.2.1. Сенсорные рецепторы Сенсорные рецепторы в зависимости от их структурной организации и функции могут быть первично или вторично чувствующими (рис. 3.5). Первично чувствующие рецепторы — это нервные окончания отростков чувствительных нейронов. Они имеются в коже и слизистых оболочках, скелетных мышцах, сухожилиях и надкостнице, а также барьерных структурах внутренней среды ■— стенках кровеносных и лимфатических сосудов, интерстициальном пространстве, в оболочках головного и спинного мозга, ликворной системе. По характеру воспринимаемых раздражителей первично чувствующие рецепторы делят на механорецепторы (восприятие растяжения или сдавления, линейного или радиального сдвига ткани), хеморецепторы (восприятие химических раздражителей), терморецепторы (восприятие температуры). В особую группу выделяют ноцицепторы, т. е. рецепторы, воспринимающие боль, хотя их существование признается не всеми. Вторично чувствующие рецепторы — это специализированные на восприятии определенных раздражителей рецепторные клетки, как правило эпителиальной природы, входящие в состав органов чувств — зрения, слуха, вкуса, равновесия. После восприятия раздражителя эти рецепторные клетки передают информацию на окончания афферентных проводников чувствительных нейронов. Таким образом, афферентные нейроны нервной системы получают информацию о раздражителе, уже обработанную в рецепторных клетках (что и определило название этих рецепторов). Все виды рецепторов в зависимости от источника воспринимаемого раздражения делят на экстероцепторы (воспринимающие стимулы из внешней среды) и интероцепторы (предназначенные для раздражителей внутренней среды). Среди интероцепторов выделяют проприоцепторы, т. е. собственные рецепторы опорно-двигательного аппарата, ангиорецепторы, расположенные в стенках сосудов, и тканевые рецепторы, локализованные в интерстициальном пространстве и клеточной микросреде. Осочувствительность для таких раздражителей оказывается наибольшей. Величина минимальной силы раздражителя, способной вызвать возбуждение рецептора, носит название абсолютного порога раздражения. В связи с этим раздражители, для которых рецептор обладает наивысшей чувствительностью, т. е. минимальной величиной порога, получили название адекватных. В то же время некоторые рецепторы могут реагировать и на несоответствующие их специализации раздражители (например, рецепторы органа зрения на механическое раздражение), порог для таких раздражителей, называемых неадекватными, оказывается очень высоким и требуется значительная сила раздражителя для возбуждения рецептора («искры из глаз» при ударе).

Если первично чувствующие рецепторы образованы разветвлениями отростка одного сенсорного нейрона, они формируют рецептивное поле сенсорного нейрона. Обычно рецепторы образуют в тканях скопления различной плотности. В случаях, когда эти скопления дают начало определенному рефлексу, их называют рецептивными полями рефлекса. Если в скоплении представлены разные по характеру раздражителей рецепторы, дающие начало разным рефлексам, их называют рефлексогенными зонами. Примером являются сосудистые рефлексогенные зоны (дуги аорты, каротидного синуса), где расположены и механо- и хеморецепторы, раздражение которых вызывает различные рефлекторные реакции сердечно-сосудистой, дыхательной и других систем организма. 3.2.2. Афферентные и эфферентные нервные проводники Основной функцией нервов является проведение сигналов к нервному центру от рецепторов (афферентные проводники) или от нервного центра к эффектору (эфферентные проводники). Собственно проводниками являются нервные волокна, входящие в состав периферических нервов или белого вещества головного и спинного мозга. В зависимости от диаметра, наличия или отсутствия миелиновой оболочки и скорости проведения нервных импульсов нервные волокна делят на три класса: А (четырех типов), В и С (табл. 3.1). Миелинизированные волокна более толстые, скорость проведения сигналов по ним существенно больше. Так, волокна A-типов обеспечивают проведение сигналов при Таблица 3.1. Характеристика нервных волокон периферических нервов
рефлекторной регуляции скелетных мышц. Немиелинизированные тонкие волокна С-типа участвуют в проведении сигналов, регулирующих деятельность внутренних органов, скорость проведения сигнала по ним — самая низкая.
3.2.3. Возбуждение и торможение в рефлекторной дуге При огромном количестве раздражителей, действующих одновременно на многочисленные рецепторные образования организма, наличии множества взаимосвязанных информационных каналов, в виде рефлекторных ответов реализуются лишь некоторые из воздействий. Целесообразность такого ограничения очевидна, поскольку в противном случае одновременно осуществляемые противодействующие рефлексы сделали бы просто невозможной не только регуляцию, но и саму жизнедеятельность. Поэтому, наряду с процессом возбуждения, т. е. возникновения активной деятельности, распространение которого лежит в основе всех рефлексов, существует второй процесс, ограничивающий и подавляющий возникновение и распространение возбуждения в элементах нервной системы и тем самым не позволяющий реализовываться рефлекторным актам. Этот второй основной процесс в нервной системе называется торможение. Под торможением понимают активный нервный процесс, возникающий при действии распространяющихся нервных импульсов и проявляющийся в ослаблении или подавлении возбуждения. Процесс торможения не способен распространяться, он возникает и проявляется локально. 3.2.4. Механизмы связи между звеньями рефлекторной дуги Передача возбуждения с отростка одной нервной клетки на отросток или тело другой нервной клетки возможна двумя способами: электрическим (электротоническим) и химическим. Электрический способ передачи возбуждения осуществляется благодаря тесным контактам передающей и воспринимающей структур (щель между мембранами меньше 2 нм). Передача возбуждения в таком случае осуществляется аналогично его проведению по нервным волокнам с помощью местных токов. Электрическая передача возбуждения осуществляется с высокой скоростью, близкой скорости проведения возбуждения по нервным волокнам и, так же как нерв, практически неутомляема. Основным способом передачи информации между нервными клетками является химический, реализуемый с помощью специальных образований, получивших название синапсов. Синапсы — это специализированная форма контакта между отростками нейронов и любыми возбудимыми образованиями (нейронами, мышечными или секреторными клетками), обеспечивающая передачу сигнала с помощью молекул химических веществ, называемых медиаторами. В нервной системе находятся центральные синапсы, которые образуются между отростками разных нейронов, а также между отростками и телами клеток. Соответственно их называют аксо-аксо- нальными, аксодендритными, аксосоматическими, дендросоматическими, дендро-дендритными. Количество синапсов на нейроне очень большое и достигает нескольких тысяч (рис. 3.6). В качестве примера может быть рассмотрен аксосоматический синапс (между аксоном одной нервной клетки и телом другой), структура которого показана на рис. 3.7. Аксон, подходя к телу другого нейрона, образует расширение, называемое пресинаптическим окончанием, или терминалью. Мембрана такого окончания называется пре-
Передача информации в синапсах осуществляется с помощью молекул специальных химических веществ — медиаторов, т. е. посредников передачи, образуемых в терминали и выводимых через пресинаптическую мембрану в синаптическую щель. Медиатор содержится в пресинаптической терминали в пузырьках и его выход происходит порциями (квантами), равными содержимому каждого пузырька.
Рис. 3.8. Структура нервно-мышечного синапса. 1 — пресинаптическая концевая пластинка, 2 — синаптические пузырьки с медиатором, 3 — митохондрии, 4 — синаптическая щель с квантами медиатора, 5 — складчатая постсинаптическая мембрана, 6 — рецепторы постсинаптической мембраны, 7 — саркоплазматический ретикулум, 8 — сократительный аппарат (миофибриллы) мышечной клетки.
щимися у рецепторов постсинаптической мембраны, и диффузии медиатора из синапсов в микроокружение клетки. Энергетическое и субстратно-ферментативное обеспечение синаптической передачи сигналов осуществляется с помощью аксонного транспорта из тела клетки в пресинаптическое окончание белков и ферментов, органелл, в частности митохондрий, низкомолекулярных веществ и уже синтезированных медиаторных веществ. Транспорт происходит с помощью нейрофибрилл или микротрубочек, тянущихся внутри аксона по всей его длине. Аксонный транспорт требует обязательного участия ионов кальция, а энергия для его реализации черпается из непрерывно ресинтезируемой в аксоне АТФ. Помимо транспорта веществ по аксонам из тела нейрона к синаптическому окончанию, существует и ретроградный аксонный транспорт, обеспечивающий поступление от пресинаптического окончания в тело клетки веществ, регулирующих в ней синтез белка. Периферические синапсы. Периферические синапсы расположены между окончаниями эфферентного нерва и мембраной эффектора. Для соматических рефлексов это синапсы между двигательным нервом и скелетной мышцей, поэтому их еще называют нервно-мышечными синапсами (рис. 3.8). Благодаря форме синаптического образования они получили название концевых пластинок. Строение и основные свойства концевых пластинок в общих чертах подобны центральным синапсам, однако постсинаптическая мембрана у них имеет многочисленные складки, существенно увеличивающие поверхность взаимодействия медиатора с рецепторами. 3.2.5. Нервные центры и их свойства Нервным центром называют функционально связанную совокупность нейронов, расположенных в одной или нескольких структурах центральной нервной системы и обеспечивающих регуляцию определенных функций организма. В более узком понимании применительно к рассматриваемой структуре рефлекторного акта нервный центр как аппарат управления представляет собой функциональное объединение разных нейронов, обеспечивающее реализацию конкретного рефлекса. Основные общие свойства нервных центров определяются тремя главными факторами: 1) свойствами нервных клеток, входящих в состав центра, 2) особенностями структурнофункциональных связей нейронов, 3) свойствами синапсов между нейронами центра. Функциональные свойства нервных клеток зависят от их строения, локализации и связей, устанавливаемых отростками. С позиций функционального предназначения различают 3 типа нейронов: афферентные, или чувствительные, эфферентные и интернейроны (промежуточные). Афферентные нейроны не входят в состав центра, по количеству отростков они обычно униполярны, биполярны или псевдоуниполярны (т. е. имеют один Т-образно делящийся отросток). Эти клетки получают сенсорную информацию от рецепторов и осуществляют ее передачу нейронам центра. Эфферентные нейроны обычно мультиполярны, т. е. имеют один аксон и несколько дендритов, они осуществляют передачу информации из нервного центра к эффекторам, например, мышцам. Наибольшую по количеству группу нервных клеток в центральной нервной системе составляют интернейроны, или ассоциативные нейроны, которые по своей форме обычно мультиполярны, причем отростки клеток не покидают пределов центральной нервной системы и их основной функцией является обеспечение взаимосвязей между разными группами нейронов. Интернейроны могут быть возбуждающими или тормозными, в зависимости от выделяемых на окончаниях их отростков медиаторов. Тело и дендриты нервных клеток являются структурами, которыми осуществляется интеграция поступающих к нейрону многочисленных сигналов. Функция нервных клеток связана с особенностями их метаболизма, в частности, с высоким уровнем энергетических и пластических процессов. Наиболее существенным отличием нейронов от других видов клеток организма является интенсивный обмен нуклеиновых кислот и очень высокий уровень транскрипции, т. е. синтеза молекул информационной РНК на матрицах ДНК генома. Интенсивность транскрипции в нейронах превышает более чем в 5 раз ее уровень в других клетках организма. Имеется четкая взаимосвязь уровня синтеза белка в нейронах и характера их электрической активности. Образуемые в нейронах специфические белки и пептиды участвуют в регуляции эффективности синаптической передачи сигналов и хранении информации, и, в конечном счете, реализации рефлекторных актов и особенностей поведения. Схематизировано наиболее простой формой связи нейронов центра является нервная цепь (рис. 3.9, А), в которой возбуждение последовательно
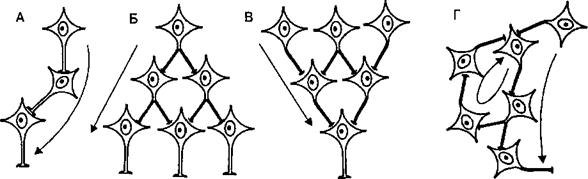
Рис. 3.9. Особенности распространения возбуждения в нейронных сетях. А — нервная цепочка, Б — дивергенция возбуждения в нервной сети, В — конвергенция возбуждения, Г — реверберация возбуждения в кольцевой цепи Лоренто де Но. Стрелки показывают направление распространения возбуждения.
передается от одного нейрона к другому. Нервная цепь может содержать всего два нейрона с одним синапсом между ними, соответственно рефлекс, реализуемый через такой нервный центр, носит название моносинап- тического. С увеличением числа нейронов и синапсов между ними время рефлекторной реакции на раздражитель увеличивается, поскольку каждый синапс имеет синаптическую задержку проведения. Такие рефлексы называются полисинаптическими. Сигналы по нервным цепям распространяются в одну сторону — от афферентного входа к эфферентному выходу — благодаря одностороннему проведению в синапсах. Нейроны нервного центра за счет структурно-функциональных связей (ветвления отростков и установления множества синапсов между разными клетками) объединяются в нервные сети. Связи между нервными клетками являются генетически детерминированными. Различают 3 основных типа нервных сетей: иерархические, локальные и дивергентные с одним входом. Иерархические сети обеспечивают постепенное включение нейронных структур более высокого уровня. В тех случаях, когда афферентная информация поступает к увеличивающемуся числу нейронов, принято говорить о дивергенции возбуждения (рис. 3.9, Б). Если, напротив, от многих возбужденных клеток информация (возбуждение) сходится к меньшему числу нейронов, то такой принцип распространения сигналов называется конвергенцией (рис. 3.9, В). Так, например, конвергенция в эффекторном пути двигательных спинальных рефлексов ведет к тому, что малое число двигательных клеток (мотонейронов) спинного мозга получает импульсы возбуждения от различных эфферентных путей многих рефлекторных дуг. Изучая этот механизм на уровне спинного мозга, Ч. Шеррингтон сформулировал принцип общего конечного пути, согласно которому мотонейроны спинного мозга являются общим конечным путем многочисленных рефлексов (мотонейроны, управляющие сгибателями правой руки, участвуют в многочисленных двигательных рефлекторных реакциях — почесывании, жестикуляции при речи, переносе пищи в рот и т. п.). Соответственно на уровне многочисленных синапсов конвергентных путей возникает конкуренция за общий конечный путь. Однако конвергенция существует и для афферентной информации, когда на ограниченном числе нейронов конвергируют сигналы, приходящие от разных сенсорных систем (слуха, обоняния, зрения и т. п.), что позволяет формировать в нервных центрах полноценный образ воспринимаемого объекта. Нервные сети обеспечивают реализацию принципа субординации, когда деятельность ниже расположенных нейронных структур подчинена выше расположенным. «Выше» и «ниже» здесь означает уровень расположения в отделах мозга, например, нейроны спинного мозга подчинены влияниям со стороны головного мозга. Локальные сети нервных центров содержат нейроны с короткими аксонами, осуществляющие взаимосвязи в пределах одного уровня. Они обеспечивают задержку информации в пределах этого уровня, что играет роль в механизмах памяти. Примером такой локальной сети являются кольцевые нейронные цепочки Лоренто де Но, возбуждение в которых циркулирует по замкнутому кругу (рис. 3.9, Г). Возврат возбуждения к «первому»нейро- ну кольцевой цепи получил название реверберации возбуждения. Локальные сети обеспечивают надежность нервной регуляции за счет дублирования элементов, так как многие нейроны локальных сетей имеют одинаковые синаптические связи и функционируют попеременно, т. е. являются взаимозаменяемыми. Дивергентные сети с одним входом представляют собой нейронные ансамбли, в которых один нейрон образует выходные связи с большим количеством других клеток разных иерархических уровней и, главное, разных нервных центров. Максимально выраженная дивергенция связей разных нервных центров свидетельствует о том, что эти нервные сети не являются специфическими для реализации определенных рефлексов, а обеспечивают интеграцию разных рефлекторных актов и общее состояние активности многочисленных нейронов разных отделов мозга. Поскольку в нервных сетях включено огромное число интернейронов, они могут, передавая нервные импульсы другим нейронам центра, реализовывать как их возбуждение, так и торможение. Различают следующие типы тормозных процессов в нейронных сетях: • реципрокное торможение (рис. 3.10, В), т. е. процесс, основанный на том, что сигналы по одним и тем же афферентным путям обеспечивают возбуждение одной группы нейронов, а через вставочные тормозные клетки вызывают торможение другой группы нейронов. Этот тип торможения называют иначе сопряженным. Проявляется реципрокное торможение, например, на уровне мотонейронов спинного мозга, иннервирующих мышцы-антагонисты (сгибатели-разгибатели) конечностей. Передача возбуждения к мотонейронам мышц-сгибателей одновременно через вставочные тормозные нейроны тормозит мотонейроны мышц-разгибателей, благодаря чему только и возможно осуществление сгибательного рефлекса; • возвратное торможение (рис. 3.10, Г), т. е. процесс торможения эфферентных нейронов сигналами, возвращающимися к ним по коллатералям аксонов через вставочные тормозные клетки. Возбуждение от одних нейронов поступает через ответвления на аксонах к вставочным тормозным нейронам, а аксоны этих клеток в свою очередь образуют тормозные синапсы на телах первых нейронов. Примером является торможение Реншоу в спинном мозге, где импульсы от двигательных нейронов посылаются по аксонам не только к скелетной мышце, но поступают через аксонные коллатерали к тормозным интернейронам Реншоу, а от них назад к двигательным нейронам, приводя к их торможению. Таким образом осуществляется короткая отрицательная обратная связь, не позволяющая возникнуть избыточному возбуждению мотонейронов спинного мозга; • латеральное торможение (рис. 3.10, Б), т. е. процесс торможения
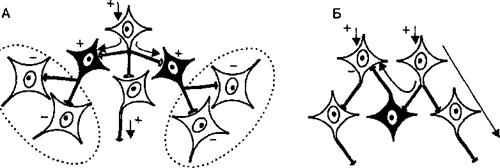
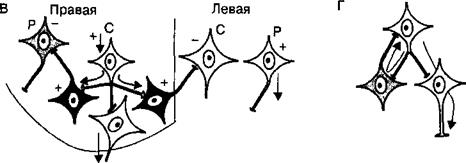
Рис. 3.10. Тормозные процессы в нейронных сетях центра. А — окружающая тормозная зона, Б — латеральное торможение, В — реципрокное торможение в центрах спинного мозга, управляющих сокращениями мышц-сгибателей (С) и -разгибателей (Р) нижних конечностей, Г — возвратное торможение (Реншоу) в спинном мозге. Знаки (+) и (-) — соответственно возбужденное и заторможенное состояние нейронов, стрелки — направление распространения возбуждения.
группы нейронов, расположенной рядом с группой возбужденных клеток. Этот вид торможения распространен в сенсорных системах, где ответвления аксонов возбужденных нейронов образуют синапсы на вставочных тормозных нейронах, а аксоны последних оканчиваются тормозными синапсами на соседних, рядом расположенных клетках, получающих информацию от другого афферентного входа. Благодаря явлению латерального торможения удается различать контраст в освещенности объектов, звуковые тоны и другие различия воспринимаемых раздражителей; • тормозная зона, т. е. процесс торможения нескольких групп нейронов, расположенных рядом с возбужденными клетками. Сигналы от возбужденных нейронов по коллатералям аксонов поступают к вставочным нейронам, а аксоны последних образуют тормозные синапсы на группах клеток, получающих информацию по другим афферентным входам (рис. 3.10, А). Если эта группа заторможенных клеток располагается вокруг возбужденных нейронов, тормозную зону называют окружающей; • нисходящее торможение, т. е. процесс торможения ниже расположенных или эволюционно более древних структур нервной системы выше расположенными или эволюционно более новыми нейронными структурами головного мозга. Реализация нисходящего торможения способствует реализации принципа субординации нервных центров. 3.2.6. Взаимодействие различных рефлексов. Принципы координации рефлекторной деятельности Поскольку в организме одновременно или последовательно реализуется обычно несколько рефлексов, простейшие связи между ними характеризуются тремя вариантами взаимодействия. Во-первых, рефлексы могут взаимно содействовать друг другу, облегчая реализацию требуемого эффекта. Такие рефлексы называют аллиированными, или союзными. Примерами ал- лиированных рефлексов являются рефлексы мигания и слезоотделения, слюноотделения и глотания. Во-вторых, рефлексы могут оказывать друг на друга тормозящее влияние, и тогда их называют антагонистическими. Например, рефлекс глотания тормозит рефлекс вдоха, рефлекс разгибания тормозит рефлекс сгибания конечности. В-третьих, взаимодействие между рефлексами может быть последовательным, когда один рефлекс, а вернее его результат, вызывает возникновение другого рефлекса. Такие рефлексы называют цепными, причем в цепи может быть взаимосвязано большое число разных рефлексов. Простейшими цепными рефлексами являются, например, шагательный рефлекс, когда сгибательный рефлекс (сгибание ноги) становится причиной следующего за ним разгибательного, а он в свою очередь вызывает сгибательный рефлекс и т. д. При реализации сложных рефлекторных реакций, обеспечивающих регуляцию жизнедеятельности и приспособление организма к меняющимся условиям среды, необходимым условием является координация рефлексов, в основе которой лежит согласованное формирование и взаимодействие процессов возбуждения и торможения в центральной нервной системе. Основными принципами координации являются: принцип сопряженного торможения (реципрокности), принцип доминанты, принцип общего конечного пути, принцип субординации нервных центров, принцип обратной афферентации. • Принцип сопряженного торможения, или реципрокности, базируется на описанном выше одноименном виде торможения и означает возникновение при возбуждении одного центра одновременного торможения другого центра для обеспечения цепных рефлексов и сложной функции. Примером реципрокности может быть регуляция спинным мозгом противоположных по функциональному назначению мышц конечностей (рис. 3.11). Так, при возбуждении мотонейронов, иннервирующих мышцы-сгибатели правой ноги, реципрокно тормозятся мотонейроны мышц-разгибателей этой ноги и возбуждаются мотонейроны мышц-разгибателей левой ноги. Формирующийся цепной характер рефлексов вслед за этим вызывает возбуждение мотонейронов разгибателей правой ноги и реципрокно — торможение мотонейронов сгибателей правой ноги и возбуждение мотонейронов сгибателей левой ноги. Таким образом реципрокные взаимоотношения между указанными рефлексами обеспечивают цепной шагательный рефлекс. • Принцип доминанты основан на открытом А. А. Ухтомским свойстве нервных центров. Доминантой называют общий принцип деятельности нервной системы, проявляющийся в виде господствующей в течение определенного времени системы рефлексов, реализуемых доминирующими центрами, которые подчиняют себе или подавляют деятельность других нервных центров и рефлексов. Нейроны доминини- рующих центров становятся более возбудимыми и способны эффективнее осуществлять пространственную и временную суммацию
Рис. 3.11. Принцип реципрокности в рефлекторной регуляции мышц-антагонистов. Возбуждение мотонейронов (1) мышцы-сгибателя тормозит через тормозные интернейроны (2) мотонейроны мышцы-разгибателя (3). Знаки (+) и (~) — соответственно возбуждение и торможение.
нервных импульсов. Синаптическое проведение к этим нейронам облегчено, и поэтому они могут возбуждаться и за счет «посторонних» импульсов от не имеющих прямых связей с доминирующими центрами информационных каналов. Вследствие суммации многочисленных сигналов возбуждение нейронов, как и число возбужденных клеток в доминирующем центре, нарастает и осуществляемые им рефлекторные реакции легко реализуются. Преобладание рефлексов доминирующего центра над другими рефлекторными актами становится особенно выраженным, поскольку через систему вставочных нейронов доминирующий центр сопряженно тормозит другие центры и текущие рефлексы. Принцип доминанты позволяет концентрировать внимание и строить поведение для достижения определенной намеченной цели. • Принцип общего конечного пути, описанный Ч. Шеррингтоном, рассматривался при изложении процесса конвергенции возбуждения. • Принцип субординации нервных центров (принцип подчинения) Проявляется в виде регулирующего влияния выше расположенных нервных центров на ниже расположенные. Так, двигательные центры головного мозга управляют спинальными мотонейронами. Примером такого влияния является феномен центрального торможения спинальных рефлексов, открытый И. М. Сеченовым и получивший название сеченовского торможения. В эксперименте И. М. Сеченова раздражение зрительных бугров лягушки с помощью кристаллика поваренной соли (т. е. раздражение ретикулярной формации среднего мозга) приводило к торможению спинальных двигательных рефлексов, вызываемых погружением лапки лягушки в слабый раствор кислоты. Следовательно, торможение центров спинного мозга явилось следствием возбуждения центров среднего мозга. Прекращение этого тормозного контроля при перерыве цереброспинальных проводящих путей вызывает резкое повышение возбудимости спинальных центров и гиперрефлексию. • Принцип обратной афферентации заключается в рецепторном восприятии результатов рефлекторного акта и проведении информации обратно в структуры нервного центра, где она обрабатывается и сравнивается с сохраняющимися параметрами возбуждения. Обратная аффе- рентация реализуется в виде механизмов положительной или отрицательной обратной связи. Таким образом, с помощью обратной афферентации нервные центры осуществляют непрерывный контроль эффективности, целесообразности и оптимальности рефлекторной деятельности. 3.2.7. Рефлекторная регуляция висцеральных функций Висцеральными функциями называют физиологические процессы, осуществляемые внутренними органами, железами, сердцем, кровеносными и лимфатическими сосудами, гладкой мускулатурой, клетками крови и направленные на поддержание обмена веществ, роста, развития и размножения. Регуляция висцеральных функций осуществляется вегетативной нервной системой, реализующей эфферентные сигналы рефлексов, получивших название вегетативных. Так, например, при растяжении желудка пищей происходит раздражение механорецепторов, заложенных в его стенке и являющихся окончаниями блуждающего нерва. По афферентным волокнам этого нерва импульсы от рецепторов поступают в нервный центр продолговатого мозга, а из него по эфферентным волокнам того же блуждающего Нерва посылаются к сердцу, вызывая замедление ритма его сокращений. Таким образом, структурной основой вегетативных рефлексов, как и соматических, является рефлекторная дуга (или с обратной связью — кольцо).
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2020-12-09; просмотров: 169; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.142.199.54 (0.022 с.) |



 лучающих, обрабатывающих и выдающих информацию нервных кле
лучающих, обрабатывающих и выдающих информацию нервных кле эфферентные, или двигательные, нервные проводники (каналы вы
эфферентные, или двигательные, нервные проводники (каналы вы