
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Значения корреляций между членами семей по показателям субъективного благополучия (по Nes, R.B. и др., 2010)Содержание книги
Поиск на нашем сайте
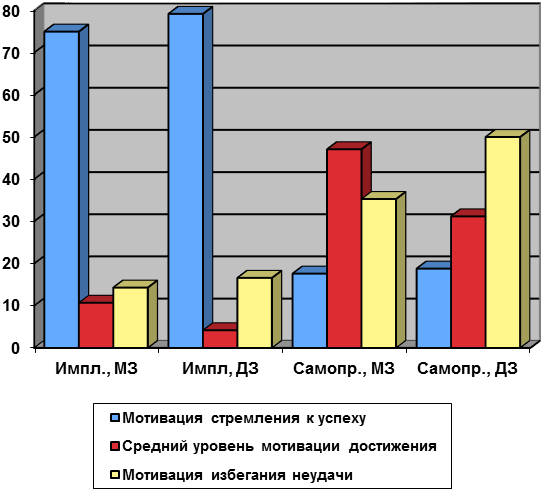
* Возраст супругов указан сначала для мужа, затем для жены. Как видно из таблицы 17 значения корреляций между членами семей по показателям субъективного благополучия статистически значимы для всех пар. Корреляции монозиготных близнецов наиболее высоки и более чем в 2 раза превышают корреляции между другими братьями и сестрами, в том числе и ДЗ близнецами. Это указывает либо на наличие влияния либо неаддитивных генетических эффектов либо индивидуальной среды на субъективное благополучие. В результате проведенного Nes, R.B. и др. (2009) исследования было получено, что оценка наследуемости субъективного благополучия в целом составила 0,33 у женщин и 0,36 у мужчин. Способы, которыми гены влияют на субъективное ощущение благополучия, пока еще неизвестны. Генетические факторы могут оказывать влияние на субъективное ощущение благополучия через эмоциональное напряжение или эмоциональную реактивность, но могут воздействовать через индивидуально-средовые факторы, а также физиологические и биохимические процессы. Нейробиологические исследования, например, показали, что устойчивость к стрессу и благополучие связаны с высоким уровнем активации левой префронтальной области коры мозга, а также активацией миндалины, что проявляется как способсность к быстрому восстановлению в ответ на негативные жизненные события (Davidson 2004). Эти области коры также связаны с общительностью у детей раннего возраста и, как было показано, участвуют в передаче дофамина, который играет важную роль в реализации положительных эмоций и экстраверсии (Depue, 1995;. Eid и др., 2003). Генетические влиния, следовательно, могут обуславливать деятельность сложных поведенческих, биохимических и физиологических систем, которые косвенно влияют на субъективное ощущение благополучия. В настоящее время поиск генов-кандидатов, связанных с ощущением субъективной удовлетворенности и чувством счастья, показали, что таким геном является, в частности, ген переносчик серотонина 5-HTTLPR. В работе де Неве и др. (2011) были использованы данные Американского национального лонгитюдного исследования здоровья подростков (которое проводилось в 1995–2008 годах). В исследовании была собрана генетическая по 2574 подросткам, а также результаты их анкетирования. В анкете, в частности, содержался вопрос о том, «Насколько вы в целом удовлетворены своей жизнью?». Респонденты должны были выбрать ответ из следующих вариантов: «весьма удовлетворен», «удовлетворен», «не удовлетворен, но и не недоволен», «недоволен» и «крайне недоволен». Де Неве с коллегами обнаружили корреляцию между ответами подростков на этот вопрос и вариацией гена 5-HTTLPR. Оказалось, что если человек имеет «длинную» вариацию этого гена, то он более подвержен влиянию серотонина и, как следствие, более счастлив. Если же у человека более «короткие» вариации гена 5-HTTLPR, тогда человек больше склонен к пессимизму (р = 0,01). В результате получено, что из 40% подростков, которые заявили, что «весьма удовлетворены» своей жизнью, 35,4% имели две «длинные» вариации «гена счастья», а 19,1% — две «короткие». В работе показано, что наличие одной «длинной» вариации гена 5-HTTLPR увеличило бы число людей, «весьма удовлетворенных» своей жизнью на 8,5%, а наличие двух «длинных» вариаций на 17,3%. В работе Song, Z. L., Li, W. D., & Arvey, R. D. (2011) было показано, что ген переносчик серотонина 5-HTTLPR связан и с удовлетворенностью супружеством, и с удовлетворенностю работой. Так, у людей, получивших от каждого из родителей по укороченному варианту гена переносчика серотонина 5-HTTLPR, обнаружена корреляция между эмоциональной тональностью общения с супругом и заявленной степенью удовлетворенности браком. Также показано, что люди с длинным аллелем гена 5-HTTLPR, как правило, больше удовлетворены работой, чем те, кто обладает коротким аллелем. Оказалось, что к генетическим маркерам удовлетворенности работой, помимо гена транспортера серотонина 5-HTTLPR относится и ген дофаминового рецептора DRD4. Известно, что дофамин и серотонин – это нейромедиаторы, играющие важную роль в «системе вознаграждения» мозга. Люди, имеющие аллель 7R гена DRD4 VNTR, как правило, удовлетворены работой в меньшей степени, чем те, кто не имеет аллель 7R. Известно, что ген DRD4 7R связан с такими характеристиками, как толерантность к риску, слабый контроль импульсов и синдром дефицита внимания и гиперактивности. Авторы предполагают, что промежуточным влиянием именно этих характеристик можно объяснить негативную связь между DRD4 7R и удовлетворённостью работой. Необходимо отметить, что влияние генов на удовлетворённость работой всегда опосредовано такими характеристиками человека, как личностные особенности, самооценка и др. Однако результаты исследований генов-маркеров удовлетворенности работой говорят о том, что руководство организации ограничено в возможностях влияния на удовлетворённость работой у своих сотрудников. Проблема исследования мотивации традиционно актуальна как в отечественной, так и в зарубежной психологии. Из существующих видов мотивации в психогенетике изучению подвергались: мотивация достижения, мотивация власти, мотивация аффилиации, мотивация агрессивного поведения. Немецкое лонгитюдное исследование близнецов (К. Готтшальдт) показало, что из трех изучаемых видов мотивации наименьшее влияние со стороны генотипа обнаруживается в отношении мотивов власти и достижений, более выраженное - в отношении мотива аффилиации (Восьмая..., 2003). Мотивация достижения – специфический вид психической регуляции деятельности, особая форма внутренней активности субъекта, представляющая собой сложную по своей природе функциональную систему интегрированных воедино аффективных и когнитивных процессов. Х. Хекхаузен определяет мотивацию достижения «как попытку увеличить или сохранить максимально высокими способности человека ко всем видам деятельности, к которым могут быть применены критерии успешности и где выполнение подобной деятельности может, следовательно, привести или к успеху, или к неудаче» (Х. Хекхаузен, 2001). Исследование мотивации достижения (М.С. Егорова, Н.М. Зырянова, О.В. Паршикова, С.Д. Пьянкова и Ю.Д. Черткова). Опросник А. Мехрабиана разработан на основе теории мотивации достижения Дж. Аткинсона и предназначен для измерения результирующей тенденции между стремлением добиться успеха и желанием избежать неудачи. В работе участвовали 65 пар МЗ и 41 пара ДЗ близнецов. Генетические факторы определяют 20% вариативности мотивации достижения. Поскольку мотивация достижения не является базовым свойством личности, то ее связь с генотипом может быть объяснена, по-мнению авторов исследования, «либо ее включением в качестве составляющей в базовые свойства личности (например, сознательность, контроль импульсивности), либо связью с генетически обусловленными когнитивными характеристиками (чем выше обобщенные показатели интеллекта, тем выше значение показателя стремления к успеху)». Исследование мотивации достижения (Е.В. Воробьева, 2009а). Проведению эмпирического исследования предшествовал теоретический анализ, в ходе которого были определены теоретические основания для проведения психогенетического исследования мотивации достижений (Е.В. Воробьева, 2003; Е.В. Воробьева, 2004; Е.В. Воробьева, 2006б; Е.В. Воробьева, 2009а). Мотивация достижения изучалась у 85 пар монозиготных и 84 пар дизиготных близнецов в возрасте 16-20 лет. Для диагностики мотивации достижения использовался опросник А. Мехрабиана и проективный тест Х. Хекхаузена. Мотивация достижения – психологический конструкт, отражающий стремление человека добиться значимых (весомых) результатов в определенной деятельности (чаще всего результаты связывают с учебной или профессиональной деятельностью). На рисунке 5 представлено частотное распределение показателей имплицитной мотивации достижения имплицитной (тестированной с помощью проективного теста Х.Хекхаузена) и самоприписываемой мотивации достижения (диагностированной с применением опросника Мехрабиана). На рисунке 5 видно, что по итогам тестирования монозиготных и дизиготных близнецов с помощью теста Х.Хекхаузена, направленного на определение уровня имплицитной (неосознаваемой) мотивации было получено, что наиболее выраженным как у монозиготных (75%), так и у дизиготных (79,2%) близнецов является уровень «стремление к успеху», что в свою очередь и определяет ведущий мотив подавляющего большинства испытуемых. Избегание неуспеха у монозиготных (14,3%) и дизиготных (16,6%) близнецов находится примерно в таком же диапазоне, как и «средний уровень», отмеченный на шкале мотивации достижений. Полученные данные оценки самоприписываемой мотивации достижения распределились по шкале мотивации достижений в диапазоне от линии «стремления к успеху», до линии «избегания неудач». Уровень «стремления к успеху» является наименее выраженным в равной степени как у монозиготных (17,6%), так и у дизиготных (18,8%) близнецов. «Средний уровень» существенно преобладает
Рисунок 5. Частотное распределение показателей имплицитной (импл.) и самоприписываемой (самопр.) мотивации достижения у монозиготных (МЗ) и дизиготных близнецов (ДЗ) (в %) (Е.В. Воробьева, 2009а)
у монозиготных близнецов (47,1%), и менее выражен и дизиготных (31,2%). Стратегия «избегания неудач» более характерна для дизиготных близнецов (50%), но она также находится в приоритете и у монозиготных близнецов (35,3%). Результаты оценки внутрипарного сходства МЗ и ДЗ близнецов и компонентов фенотипической дисперсии показателей мотивации достижения представлены в таблице 18.
Таблица 18 Внутрипарное сходство МЗ и ДЗ близнецов, вклад генетической (h²), общесемейной (с²) и индивидуально-средовой (е²) составляющих фенотипической дисперсии мотивации достижения, диагностированной тестами Х. Хекхаузена и А. Мехрабиана (*p<0,05; **p<0,01) (Е.В. Воробьева, 2009а)
В таблице 18 представлены данные, свидетельствующие о том, что наибольшее влияние на фенотипическую вариативность мотивации достижения оказывают индивидуально-средовые факторы, кроме того, для мотивации достижения, оцениваемой с применением опросника, установлено наличие и достаточно выраженного влияния генетических факторов. В своих последующих работах (Ermakov P.N., Vorobyeva E.V., 2011) мы рассматривали особенности спектральных характеристик ЭЭГ у людей с выраженностью мотивации стремления к успеху или избегания неудачи, что позволило выделить и описать такие характеристики (см. Тему 5 данного учебника). В работе Д.С. Ров, Д.М. Алмейда, К.С. Якобсон были установлены особенности средовых влияний на проявление мотивации агрессивности у детей. Средовые влияния на проявление мотивации агрессивности у детей выше в группе, характеризующейся низкими средними показателями по семейному теплу и сплоченности. Наследуемость мотивации агрессивности выше в группе, где выше показатели семейного тепла и сплоченности. Сравнение оценок внутрипарного сходства у МЗ и ДЗ близнецов позволяет сделать выводы о наследуемости мотивации агрессии (Rowe D.C., 1999).
Список литературы к теме 8:
1. Айзенк Г.Ю. Как измерить личность. М.: Когито-Центр. 2000. 284 с. 2. Алфимова М.В., Трубников В.И Психогенетика агрессивности // Вопросы психологии. № 6. 2000. C. 112-121. 3. Богомолова М.В. Влияние обогащенной среды на развитие интеллекта и креативности. Автореферат диссертации на соискание ученой степени кандидата психологических наук. Москва ИП РАН, 2008. 22 с. 4. Бэрон Р., Ричардсон Д. Агрессия. — СПб: Питер, 2001. — 352 с: 5. Воробьева Е.В. Взаимосвязь параметров общего интеллекта и мотивации достижения // Северо-Кавказский психологический вестник. 2009(а). № 7/3. С. 25-33. 6. Воробьева Е.В. Интеллект и мотивация достижения: психогенетический аспект // Валеология. 2003. № 4. С. 46-51. 7. Воробьева Е.В. Психогенетическое исследование взаимосвязи темперамента и мотивации достижения // Известия высших учебных заведений. Северо-Кавказский регион. Общественные науки. 2006(а). № 3. С. 110-111. 8. Воробьева Е.В. Современные психогенетические исследования интеллекта и теория мотивации достижения // Журнал прикладной психологии. 2004. № 3. С. 53-59. (0,55 п.л.). 9. Воробьева Е.В. Функционально-регуляционная модель взаимосвязи интеллекта и мотивации достижения // Северо-Кавказский психологический вестник. 2006(б). № 4. С. 172-179. 10. Воробьева Е.В., Нарижная Е.С. Особенности темперамента студентов-психологов, различающихся по мотивации достижения / Психологический институт в современном научно-психологическом пространстве: Международные Челпановские чтения 2014: Московская научно-практическая конференция к 100-летию Торжественного открытия Психологического института им. Л. Г. Щукиной (1914–2014). Москва, 22–23 апреля 2014года / Альманах Научного архива Психологического института / Сост., научн. ред. О. Е. Серова, Е. П. Гусева / Под общ. ред. В. В. Рубцова. М.: Алькор Паблишер, 2014. Вып. 7 юбилейный. – С. 299-306. 11. Воробьёва Е.В., Попова В.А. Исследование интеллекта и мотивации достижения близнецов // Российский психологический журнал. 2009(б). Т.6. С. 46-53. 12. Воробьева Е.В., Харитонова И.Ю. Генотип-средовые детерминанты мощности ритмических составляющих ЭЭГ при вербально-ассоциативной деятельности // Новые исследования. М.: Вердана. 2010. T.1 № 23. С.5-16. 13. Дружинин В.Н. Психология общих способностей. 2-е издание. С.-Пб.: «Питер», 1999. 368 с. 14. Дружинин В.Н., Воробьева Е.В. Воздействие общения экспериментатора с испытуемым на проявление психометрического интеллекта у подростков-монозиготных близнецов // Психологический журнал. 1997. т. 18. № 1. С. 70-80. 15. Егорова М.С., Зырянова Н.М., Паршикова О.В., Пьянкова С.Д., Черткова Ю.Д. Генотип. Среда. Развитие. М.: ОГИ, 2004. 576 с. 16. Егорова М.С., Зырянова Н.М., Пьянкова С.Д. Возрастные изменения генотип-средовых соотношений в показателях интеллекта // Вопросы психологии. 1993. № 5. С. 106-108. 17. Ильин Е.П. Дифференциальная психофизиология. С.-Пб.: Питер, 2001. 454 с. 18. Кантонистова Н.С. Роль гено и паратипических факторов в формировании отдельных сторон интеллектуальной деятельности: Автореф. канд. дисс. М., 1978. 22 с. 19. Новгородова Ю.О., Мухордова О.Е., Сивакова В.Г., Лобаскова М.М., Барский Ф.И., Малых С.Б. Генетические и средовые факторы в формировании индивидуальных особенностей темперамента у детей младшего школьного возраста // Теоретическая и экспериментальная психология. 2010. Т. 3. 20. Новые тесты IQ. 3-е издание. «Феникс», 2004. 352 с. 21. Малых С.Б. Психогенетика: теория, методология, эксперимент. М.: Эпидавр, 2004. 416 с. 22. Малых С.Б., Гиндина Е.Д., Надысева В.В. Природа индивидуальных особенностей темперамента в подростковом возрасте // Психологический журнал. 2004. т. 25. № 6. С. 29-52. 23. Малых С.Б., Егорова М.С., Мешкова Т.А. Основы психогенетики. М., 1998. 24. Практикум по психологии менеджмента и профессиональной деятельности / Под ред. Г.С. Никифорова, М.А. Дмитриевой, В.М. Снеткова. СПб.: Речь, 2001. 448 с. 25. Равич-Щербо И.В., Марютина Т.М., Григоренко Е.Л. Психогенетика. М.: Аспект-пресс, 1999. 447 с. 26. Русалов В.М., Русалова М.Н., Стрельникова Е.В. Психофизиологические предпосылки предпочтения высокой вероятности достижения цели или ее субъективной ценности // Физиология человека. 2000. Т. 26. № 5. С. 69. 27. Русалов В.М., Русалова М.Н., Стрельникова Е.В. 28. Русалов В.М., Трофимова И.Н. О представленности типов психической деятельности в различных моделях темперамента / Психологический журнал. № 3. 2011. С. 74 -84. 29. Сергиенко Е.А., Рязанова Т.Б. Младенческое близнецовое исследование: специфика психического развития // Психол. журнал. т. 20. № 2. 1999. С. 39-53. 30. Собчик Л.Н. Мотивационный тест Х. Хекхаузена. Практическое руководство. СПб.: Речь, 2002. 16 с. 31. Ушаков Д.В. Интеллект: структурно-динамическая теория. М.: ИП РАН, 2003. 263 с. 32. Ando J., Y. Ono, M.J. Wright Genetic Structure of Spatial and Verbal Working Memory // Behavior Genetics. 2001. V. 31. №. 6. P. 615-624. 33. Bartels M, Boomsma D (2009) Born to be happy? The etiology of subjective well-being. Behav Genet. Published online 03 September 2009. 34. Bartels M., Reitveld M.J.H., van Baal G.C.M., Boovsma D.I. Genetic and Environmental Influences on the Development of Intelligence // Behavior Genetics. 2002. V. 32, N. 4. P. 237-249. 35. Carroll, J.B. (1993). Human cognitive abilities: A survey of factor-analytic studies. Cambridge, England: Cambridge University Press. 36. Caprara GV, Fagnani C, Alessandri G, Steca P, Gigantesco A, Cavalli-Sforza L, Stazi MA (2009) Human optimal functioning. The genetics of positive orientation towards self, life, and the future. Behav Genet 39:277–284. 37. Celec P., Ostatníková D., Holešová Z., Minárik G., Ficek A., Kelemenová S., Putz Z., Kúdela M. Spatial Abilities in Prepubertal Intellectually Gifted Boys and Genetic Polymorphisms Related to Testosterone Metabolism // Journal of Psychophysiology. 2009. V. 23. Issue 2. P. 1-6. 38. Christensen K., Frederiksen H., Vaupel J. W., and McGue M. Age Trajectories of Genetic Variance in Physical Functioning: A Longitudional Study of Danish Twins Aged 70 Years and Older // Behavior Genetics. 2003. V. 33. № 2. P. 125-136. 39. Davidson R.J. (2004) Well-being and affective style: neural substrates and bio-behavioural correlates. Philos Trans R Soc Lond B 359:1395–1411. 40. De Neve J.-E. Functional polymorphism (5-HTTLPR) in the serotonin transporter gene is associated with subjective well-being: evidence from a US nationally representative sample / Journal of Human Genetics. June 2011. 56. P. 456-459. 41. Deary Ian J., Steve Strand, Pauline Smith, Cres Fernandes Intelligence and educational achievement // Intelligence. 2007. V. 35. P. 13–21. 42. Deary I., Whalley L.J., Clair D.S., Breen G., Leaper S., Lemmon H., Hayward C., Starr J. M. The influence of the ε4 allele of the apoliprotein E gene on childhood IQ, nonverbal reasoning in old age, and lifetime cognitive change // Intelligence. 2003. 31. P. 85-92. 43. Depue R.A. (1995) Neurobiological factors in personality and depression. Eur J Pers 9:413–439. 44. Jacobs N., Van Gestel S., Derom C., Thiery E., Vernon P., Derom R., Vlietinck R. Heritability estimates of intelligense twins: effect of chorion type // Bеhavior Genetics. 2001. V. 31. № 2. P. 209-217. 45. Eid M., Riemann R., Angleitner A., Borkenau P. (2003) Sociability and positive emotionality: genetic and environmental contributions to the covariation between different facets of extraversion. J Pers 71:319–346. 46. Ermakov P.N., Vorobyeva E.V. Achievement Motivation and EEG Spectral Power // Psychology in Russia: State of the Art. Scientific Yearbook / Ed. By Yu. P. Zinchenko and V. F. Petrenko. – Moscow: Lomonosov State University; Russian Psychological Society, 2011. P. 448-464. 47. Fagan Joseph F., Cynthia R. Holland, Karyn Wheeler The prediction, from infancy, of adult IQ and achievement // Intelligence. 2007. V. 35. P. 225–231. 48. Johnson W., Thomas J. Bouchard Jr., Matt McGue, Nancy L. Segal, Auke Tellegen, Margaret Keyes, Irving I. Gottesman Genetic and environmental influences on the Verbal-Perceptual-Image Rotation (VPR) model of the structure of mental abilities in the Minnesota study of twins reared apart // Intelligence. 2007. V. 35. P. 542–562. 49. Haworth C.M.A., P.S. Dale, R. Plomin Generalist Genes and High Cognitive Abilities // Behav. Genet.- 2009.- № 39.- Р. 437–445. 50. Kahneman D (1994) New challenges to the rationality assumption. J Inst Theor Econ 150:18–36. 51. Larsen L., Peter Hartmann, Helmuth Nyborg The stability of general intelligence from early adulthood to middle-age // Intelligence. 2008. V. 36. P. 29–34. 52. Loehlin T.C., Nichols R.C. Heridity, Environment, and Personality: A Study of 850 Twins. Agustin, 1976. 53. Lucas R.E., Diener E. (2008) Personality and subjective well-being. In: John OP, Robins R, Pervin LA (eds) Handbook of personality, 2nd edn. Guilford, New York. 54. Lykken D., Tellegen A. (1996) Happiness is a stochastic phenomenon. Psychol Sci 7:186–189. 55. Luciano M., M.J. Wright, G.M.Geffen, L.B.Geffen, G.A. Smith, Evans D.M., N.G. Martin A genetic two-factor model of the covariation among a subset of Multidimensional Aptitude Battery and Wechsler Adult Intelligence Scale-Revise subtests // Intelligence. 2003. V. 31. P. 589-605. 56. Lyubomirsky S., King L.A., Diener E. (2005) The benefits of frequent positive affect: does happiness lead to success. Psychol Bull 131:803–855. 57. Malykh S.B., Gindina E.D., Nadyseva V.V. Genetic and environmental influences on temperament in adolescence // Psychology in Russia: State of the Art. 2009. № 2. Р. 58. McGue M., Bacon S., Lykken D.T. (1993) Personality stability and change in early adulthood: a behavioral genetic analysis. Dev Psychol 29:96–109. 59. Moum T., Næss S., Sørensen T., Tambs K., Holmen J. (1990) Hypertension labelling, life events, and psychological wellbeing. Psychol Med 20:635–646. 60. Neiss M., D. Rowe Parental Education and Child’s Verbal IQ in Adoptive and Biological Families in the National Longitudinal Study of Adolescent Healf // Behavior Genetics. 2000. V. 30. N. 6. P. 487-495. 61. Nes, R.B., Czajkowski, N., Tambs, K. Family Matters: Happiness in Nuclear Families and Twins / Behav. Genet. 2010. V. 40. P. 577–590. 62. Nes R.B., Røysamb E., Tambs K., Harris J.R., Reichborn-Kjennerud T. (2006) Subjective well-being: genetic and environmental contributions to stability and change. Psychol Med 36:1033–1042. 63. Petrill S.A., Deater-Deckard К. The heritability of general cognitive ability: A within-family adoption design // Intelligense. 2004. № 32. P. 403-409. 64. Reeve C.L., Rustin D. Meyer, Silvia Bonaccio Intelligence-personality associations reconsidered: the importance of distinguishing between general and narrow dimensions of intelligence // Intelligence. 34 (2006) Р. 387 – 402. 65. Reitveld M.J.H., G.C.M. Van Baal, C.V. Dolan, D.I. Boovsma Genetic Factor Analyses of Specific Cognitive Abilities in 5-Year-Old Dutch Childreen // Behavior Genetics. 2000. V. 30. №. 1. P. 29-40. 66. Rettew D.C., Rebollo-Mesa I., Hudziak J.J., Willemsen G., Boomsma D.I. (2008) Non-additive and additive genetic effects on extraversion in 3314 Dutch adolescent twins and their parents. Behav Genet 38:223–233. 67. Rowe D.C., David M. Almeida, and Kristen C. Jacobson Shool Contex and Genetic Influences on Aggression in Adolescence // Psychological Science. 1999. № 3. V. 10. Р. 277-280. 68. Røysamb E., Neale M.C., Tambs K., Reichborn-Kjennerud T., Harris J.R. (2003) Happiness and health: environmental and genetic contributions to the relationship between subjective well-being, perceived health, and somatic illness. J Pers Soc Psychol 85:1136–1146. 69. Song, Z.L., Li, W.D., & Arvey, R.D. (2011) Associations Between Dopamine and Serotonin Genes and Job Satisfaction: Preliminary Evidence From the Add Health Study. Journal of Applied Psychology, 96(6), 1223–1233. 70. Spinath F.M., Ronald A., Harlaar N., Price T.S., Plomin R. Phenotypic g early in life On the etiology of general cognitive ability in a large population sample of twin children aged 2-4 years // Intelligence. 2003. V. 31. P. 195-210. 71. Stubbe J.H., Posthuma D., Boomsma D.I., De Geus E.J. (2005) Heritability of life satisfaction in adults: a twin-family study. Psychol Med 35:581–1588. 72. Tambs K., Moum T. (1992) No large convergence during marriage for health, lifestyle, and personality in a large sample of Norwegian spouses. J Marriage Fam 54:957–971. 73. Tellegen A., Lykken D.T., Bouchard T.J., Wilcox K.J., Segal N.L., Rich S. (1988) Personality similarity in twins reared apart and together. J Pers Soc Psychol 54:1031–1039. 74. van der Valk J.C., van den Oord E.J., Verhulst F.C., Boomsma D.I. (2003) Genetic and environmental contributions to stability and change in children’s internalizing and externalizing problems / J. Am. Acad. Child Adolesc. Psychiatry. 42(10):1212–1220. 75. Wadsworth S.J., Corley R.P., Hewitt J.K., Plomin R., DeFries J.C. Parent-offspring resemblance for reading performance at 7, 12 and 16 years of age in Colorado Adoption Project // Journal of Child Psychology and Psychiatry. 2002. V. 43:6. P. 769-774. 76. Watson D., Clark L.A. (1997) Extraversion and its positive emotional core. In: Hogan R, Johnson J, Briggs S (eds) Handbook of personality psychology. Academic Press, New York, pp. 767–793. 77. Webbink D., Danielle Posthuma, Dorret I. Boomsma, Eco J.C. de Geus, Peter M. Visscher Do twins have lower cognitive ability than singletons? // Intelligence. 2008. V. 36. P. 539–547. 78. Weiss A., Bates T.C., Luciano M. (2008) Happiness is a personal(ity) thing: the genetics of personality and well-being in a representative sample. Psychol Sci 19:205–209.
Тема 9. Возрастная психогенетика
Краткое содержание. Возрастная психогенетика – психогенетика индивидуального развития человека. Основные модели индивидуального развития. Проблема биологического созревания и психического развития. Наследственность как фактор, типизирующий и индивидуализирующий развитие. Генетические и средовые факторы, обеспечивающие преемственность и гетерохронность развития. Смена механизмов реализации психических функций и динамика генотип-средовых соотношений в онтогенезе. Возрастная динамика генотип-средовых соотношений как основа для возрастной периодизации и для выделения сензитивных периодов развития. Р. Пломин: основные модели возрастной психогенетики.
Главным понятием возрастной психогенетики является понятие «генетическое изменение», которое характеризует изменения в эффекте действия генов на разных стадиях онтогенеза. При этом возможна оценка в разных возрастах относительной доли генетической вариативности в общей вариативности признака (что позволит оценить, как меняется наследуемость признака в ходе онтогенеза). С этой целью проводится сопоставление показателей наследуемости у аналогичных групп родственников в разных возрастах, т.е. используется вариант метода возрастных срезов, что обеспечивает выделение возрастных различий в наследуемости признаков. Также возможна и оценка того, насколько связаны между собой генетические компоненты дисперсии признака в разных возрастах. Для этого проводится лонгитюдное исследование, в котором определяется корреляция между генетическими компонентами дисперсии изучаемой характеристики, полученными в разных возрастах на одной и той же группе испытуемых. Таким образом, полная схема исследования в психогенетике развития с необходимостью включает эмпирическое исследование и наследуемости, и генетических корреляций. Сама по себе высокая наследуемость признака, полученная в разных возрастах, ни в коей мере не свидетельствует о стабильности генетических влияний. По представлениям Р. Пломина, эти две переменные — наследуемость и степень генетической общности, определяемая величиной генетической корреляции, — относительно независимы и могут образовывать разные варианты сочетаний. Модель А, например, предполагает, что наследуемость признака с возрастом может оставаться без изменений (левый столбик) или возрастать (правый), но независимо от этого генетической преемственности при этом не обнаруживается, т.е. генетические эффекты в раннем возрасте и зрелости абсолютно не связаны между собой. В моделях В и С, напротив, допускаются варианты частичной (В) или (С) полной генетически опосредованной преемственности в формировании признака. Последний вариант (с точки зрения Р. Пломина, наиболее вероятный) получил название амплификационной модели. Амплификационная модель реализации генетических влияний в онтогенезе предполагает, что с возрастом, по мере созревания ЦНС и формирования индивидуально устойчивых способов переработки информации, возрастает наследуемость признака, при этом сохраняется высокая межвозрастная генетическая корреляция, т.е. генетические эффекты, действовавшие в младенчестве, высоко коррелируют с генетическими эффектами в зрелости (Цит. по И.В. Равич-Щербо и др., 2000).
Литература к теме 9:
1. Егорова М.С., Зырянова Н.М., Паршикова О.В., Пьянкова С.Д., Черткова Ю.Д. Генотип. Среда. Развитие. М.: ОГИ, 2004. С. 576. 2. Равич-Щербо И.В., Марютина Т.М., Григоренко Е.Л. Психогенетика: Уч. для ВУЗов. М.: «Аспект Пресс», 2000. 447 с.
Задания для самоконтроля к II модулю
1. Выберите наследственно обусловленные типы ЭЭГ: а) низковольтная; б) высоковольтная; в) низковольтная пограничная; г) высоковольтная пограничная; д) затылочные медленные бета волны; е) монотонные альфа волны; ж) монотонные бета волны; з) фронто-прецентральные тета группы; и) фронто-прецентральные бета группы; к) диффузные альфа волны; л) диффузные бета волны. 2. Выберите правильное утверждение: а) наследуемость вербального интеллекта с возрастом уменьшается; б) наследуемость вербального интеллекта с возрастом остается неизменной; в) наследуемость вербального интеллекта с возрастом растет; г) динамика наследуемости вербального интеллекта с возрастом не изучена. 3. В психогенетическим иследовании вызванных потенциалов на различные стимулы, выполненном Т.М. Марютиной и Т.Г. Ивошиной (1984), максимум наследственных влияний обнаружился в реакциях: а) на семантические стимулы (рисунок дома и слово ДОМ); б) на вспышку и шахматное поле. 4. Выберите верное значение обобщенной оценки наследуемости нейротизма у взрослых: а) 20%; б) 30%; в) 40%; г) 50%; д) 60%; е) 70%. 5. Модель наследования мануальной асимметрии, предполагающая, что праворукость и локализация центров речи в левом полушарии определяется наличием гена «правостороннего сдвига», называется: а) модель М. Аннет; б) модель Дж. Леви и Т. Нагилаки; в) простая генетическая модель; г) модель И. Макмануса; д) модель К. Леланда. 6. Выберите ряд с правильно указанными оценками показателей наследуемости интеллекта в разных возрастных группах, полученные в Луисвиллском близнецовом исследовании: а) 1 год -5%; 2 года – 25%; 3 года -35%; 4 года – 45%, 15 лет – 55 %; б) 1 год -50%; 2 года – 55%; 3 года -55%; 4 года – 55%, 15 лет – 55 %; в) 1 год -55%; 2 года – 45%; 3 года -35%; 4 года – 25%, 15 лет – 20 %; г) 1 год -10%; 2 года – 17%; 3 года -18%; 4 года – 26%, 15 лет – 55 %. 7. «Генетическое изменение», как основное понятие психогенетики развития, характеризует: а) направление (генезис) основных изменений генотипа; б) изменения в составе генотипа в ходе онтогенеза; в) изменения в эффекте действия генов на разных стадиях онтогенеза. 8. Для амплитуды альфа-ритма наиболее низкие показатели наследуемости получены для отведений: а) лобных; б) затылочных; в) левых височных; г) центральных; д) правых височных. 9. Укажите метод возрастной психогенетики, который заключается в определении корреляции между генетическими компонентами дисперсии изучаемой характеристики, что позволяет судить о преемственности и стабильности генетических и средовых влияний: а) сопоставление показателей наследуемости у аналогичных групп родственников в разных возрастах (разновидность метода возрастных срезов); б) лонгитюдное исследование, проводимое в разных возрастах на одной и той же группе испытуемых (близнецов или сибсов); в) метод разлученных близнецов (разновидность метода приемных детей); г) метод анализа родословной пробанда. 10. Выберите верное значение обобщенной оценки наследуемости экстраверсии-интроверсии у взрослых: а) 20%; б) 30%; в) 40%; г) 50%; д) 60%; е) 70%. Задания для самостоятельной работы студентов к II модулю Темы для подготовки рефератов и докладов:
Литература:
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2020-11-23; просмотров: 247; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.145.156.17 (0.015 с.) |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||

 № 4.
№ 4. 


