
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Мал. 38. Основні компоненти анатомічної будови середнього мозкуСодержание книги
Поиск на нашем сайте
(рис. В.І Козлова, Т.А. Цехмистренко).
Основу середнього мозку складають ніжки мозку – два товстих білих тяжа, які ідуть від моста доверху і зовні, а потім поглинаються в речовину великого мозку. Між внутрішніми поверхнями ніжок на основі мозку знаходиться міжножкова ямка і задня продірявлена речовина. На вентральній поверхні ніжок мозку з внутрішньої сторони кожної ніжки мозку виходить окоруховий нерв (ІІІ пара). На дорсальній поверхні стовбуру позаду пластинки даху знаходиться місце виходу блокового нерву (ІV пара). Завдяки чорної речовини основа ніжок мозку відокремлена від покришки. У покришці середнього мозку під водопроводом мозку в оточуючій його сірій речовині розташовані ядра окорухового і блокового нервів, а також середньомозкове (мезенцефіличне) ядро трійчастого нерву, яке продовжується з моста. Крім цього, сіра речовина покришки середнього мозку утворює парне червоне ядро (nucl. ruber), яке є великим скупченням нервових клітин, що відіграють, поряд з іншими утвореннями, ключову роль в екстрапірамідній системі. До складу сірої речовини покришки також входять ядра ретикулярної формації.
Мал. 39. Пластинка покрівлі середнього мозку. 1-метаталамус, 2-верхні горбики, 3-нижні горбики, 4,12- блоковий н., 5-верхній мозковий парус, 6-верхня мозочкова ніжка, 7-порожнина четвертого мозкового шлуночка, 8-нижня мозочкова ніжка, 9-довгастий мозок, 10-середня мозочкова ніжка, 11-міст, 13-медіальне колінчасте тіло, 14-ніжка мозку, 15-зоровий тракт, 16-латеральне колінчасте тіло (мал. В.І. Козлова). Біла речовина покришки середнього мозку утворена різноманітними за складом волокнами. Тут проходять волокна висхідні чутливі і низхідні рухові. Висхідні сенсорні шляхи – присередня, спинномозкова і трійчаста петлі (проводять імпульси проприоцептивної і екстероцептивної чутливості) і бічна (слухова) петля – імпульси ідуть у товщі покришки ближче до її латеральної поверхні. Низхідні шляхи – покришко - спинномозковий і червоноядерно - спинномозковий – займають у покришці медіальне положення, так як вони утворюють перехрестя по серединній лінії. Покришко-спинномозковий шлях починається від ядер верхніх і нижніх горбиків пластинки покрівлі. Аксони нейронів, розташованих у цих ядрах, огинають знизу водопровід мозку і переходять на протилежний бік, утворюючи дорсальне перехрестя покришки. Після перехрестя волокна направляються у складі покришки стовбуру до передніх канатиків спинного мозку.
Червоноядерно - спинномозковий шлях починається від нейронів червоних ядер, аксони яких утворюють вентральне перехрестя покришки, після чого також у покришці стовбуру мозку направляються до бічних канатиків спинного мозку. У покришці середнього мозку, як і в інших відділах стовбура мозку, розташовані ядерні і волокнисті структури ретикулярної формації. В основі ніжок мозку проходять низхідні кірково-мосто-мозочкові і пірамідні (кірково-спинномозкові і кірково-ядерні) шляхи, які з’єднують кору головного мозку з мозочком і спинним мозком.
ПРОМІЖНИЙ МОЗОК
Проміжний мозок (diencephalon) розміщується між кінцевим і середнім мозком. На основі мозку його межа спереду проходить по передній поверхні перехрестя зорових нервів, передньому краю задньої продірявленої речовини і зоровим трактам, а позаду – по краю ніжок мозку. На дорсальній поверхні передньою межею є термінальна смужка, яка відокремлює проміжний мозок від кінцевого мозку, а задньою межею є борозна, що відокремлює проміжний мозок від верхніх горбиків середнього мозку. На сагітальному зрізі проміжний мозок розташований під мозолистим тілом і склепінням. Дорсальний відділ проміжного мозку представляє собою філогенетично більш молодий таламічний мозок, який є вищим підкорковим сенсорним центром, в якому переключаються практично всі аферентні шляхи, що несуть сенсорну інформацію від органів тіла і органів чуття. Вентральний відділ,гіпоталамус, більш старе у філогенетичному відношенні утворення, відіграє роль вищого центру регуляції вегетативних функцій організму. Таламічний мозок в свою чергу поділяється на парне утворення - таламус (зоровий горб), метаталамус (заталамічна ділянка), епіталамус (надталамічна ділянка) і субталамус (підталамічна ділянка). Порожниною проміжного мозку є ІІІ шлуночок, який через правий і лівий міжшлуночкові отвори спілкується з бічними шлуночками, які розташовані усередині великих півкуль, і завдяки водопроводу мозку з’єднується з порожниною IV шлуночка мозку. У верхній стінці ІІІ шлуночка розміщується судинне сплетення, яке приймає участь поряд із сплетеннями в інших шлуночках мозку в утворенні спинномозкової рідини.
ТАЛАМУС
Таламус (thalamus) або згір’я, представляє собою утворення яйцеподібної форми, яке складається в основному із скупчення численних ядер. Таламуси утворюються за рахунок потовщення бокових стінок проміжного мозку. Спереду загострена частина таламуса утворює передній горбик, в якому розміщуються проміжні центри сенсорних (аферентних) шляхів, що ідуть від стовбура мозку до кори великих півкуль. Задня, розширена частина таламуса – подушка – містить підкорковий зоровий центр (мал. 40). Медіальні поверхні таламусів повернуті одна до одної і утворюють бокові стінки ІІІ шлуночка. Посередині вони з’єднані між собою міжталамічним зрощенням. Латеральна поверхня таламуса межує з внутрішньою капсулою – шаром білої речовини півкуль головного мозку, що складається з проекційних волокон, які з’єднують кору великих півкуль з нижче розташованими мозковими структурами. Товща сірої речовини таламуса поділена вертикальним Y - подібним прошарком (пластинкою) білої речовини на три частини – передню, медіальну і латеральну (мал. 41). Передня частина таламуса лежить усередині роздвоєної частини пластинки, медіальна і латеральна – відповідно усередині і зовні від цієї пластинки. У кожній з цих частин таламуса знаходиться декілька груп таламічних ядер. Всього в таламусі за різними даними міститься від 40 до 150 спеціалізованих ядер, які об’єднанні в 8 груп. Розглянемо основні з них.
Мал. 40. Середній і проміжний мозок (вигляд зверху): 1-мозолисте тіло, 2-прозора перегородка, 3-ніжки склепіння, 4-порожнина третього шлуночка, 5-таламус, 6-подушка таламуса, 7-пластинка середнього мозку, 8-епіфіз, 9-головка хвостатого тіла (мал. В.І. Козлова).
У передній частині таламуса розміщується вентральна група таламічних ядер. Найбільш великі з них – передньо-вентральні і передньо-присередні ядра. Вони отримують аферентні волокна у складі сосочково - таламічного шляху від сосочкових тіл – нюхового центру проміжного мозку. Від передніх ядер еферентні волокна направляються до поясної закрутки кори великих півкуль. Передня група таламічних ядер і пов’язані з нею структури є важливим компонентом лімбічної системи мозку, яка керує психоемоційною поведінкою. У медальній частині таламуса розрізняють присередньо-дорсальне ядро і групу ядер середньої лінії. Присередньо-дорсальне ядро має двосторонні зв’язки з нюховою корою лобової частки і поясної закрутки великих півкуль, мигдалеподібним тілом і передньо-присереднім ядром таламуса. Функціонально воно тісно пов’язано також з лімбічною системою і має двосторонні зв’язки з корою тім’яної, вискової і острівцевої часток мозку. Завдяки цьому присередньо-дорсальне ядро безпосередньо приймає участь у реалізації вищих психічних процесів. Його руйнування приводить до зниження занепокоєння, тривожності, напруженості, агресивності, усуненню нав’язливих думок.
Мал. 41. Топографія ядер таламуса: А-медіальна група ядер, Б-латеральна група ядер, В-передня група ядер, 1-мозкова пластинка, 2-присереднє заднє ядро, 3-інтраламінарні ядра, 4-подушка таламуса, 5-бічні задні ядра, 6-вентро-бічні ядра.
Ядра середньої лінії займають найбільш медіальне положення у таламусі і досить численні. Вони отримують аферентні волокна від гіпоталамуса, від ядер шва і голубої плями ретикулярної формації стовбуру мозку. Сюди проектується частина волокон спинномозково-таламічних шляхів у складі присередньої (медіальної) петлі. Еферентні волокна від нейронів ядер середньої лінії направляються до гіппокампу, мигдалеподібного тіла і поясної закрутки великих півкуль, які входять до складу лімбічної системи. Зв’язки з корою великих півкуль двосторонні. Це дозволяє ядрам середньої лінії грати важливу роль у процесах пробудження і активації кори великих півкуль, а також у забезпеченні процесів пам’яті. У латеральній частині таламуса розміщуються задня бічна, передня бічна, вентральна задньо-присередня і задня групи ядер. Ядра передньої бічної групи анатомічно і функціонально суттєво розрізняються між собою. Задні ядра передньої бічної групи часто розглядаються як одне вентро-бічне ядро таламуса. Ця група ядер отримує основну масу волокон висхідних шляхів загальної чутливості у складі медіальної петлі. Суди надходять також волокна смакової чутливості і волокна вестибулярних ядер. Еферентні волокна, які починаються від ядер вентро-бічної групи, направляються до кори тім’яної частки великих півкуль, куди приводять соматосенсорну інформацію від всього тіла. Задня група ядер таламуса складається з клітинних скупчень подушки таламуса. До ядер ідуть аферентні волокна від верхніх горбиків пластинки середнього мозку і волокна у складі зорових трактів. Еферентні волокна, які починаються від ядер задньої групи, широко розповсюдженні в корі лобової, тім’яної, потиличної, вискової і лімбічної часток великих півкуль. Ядерні центри подушки таламуса мають відношення до зору і причетні до комплексного аналізу різних сенсорних подразників. Вони відіграють значну роль у перцептивній (пов’язані з сприйняттям) і когнітивній (пізнавальній, розумовій) діяльності мозку, а також у зберіганні і відтворенні інформації – пам’яті. Інтраламінарна група ядер таламуса лежить у товщі вертикальної Y – подібної прошарки білої речовини (рис.41). Інтраламінарні ядра взаємопов’язані з базальними ядрами, зубчастим ядром мозочка і корою великих півкуль. У функціональному відношенні ці ядра відіграють важливу роль в активаційній системі мозку. Пошкодження інтраламінарних ядер в обох таламусах приводить до різкого зниження рухової активності (кінетичний мутізм), а також апатії і руйнуванню мотиваційної структури особи.
Слід відмітити, що кора великих півкуль завдяки двостороннім зв’язкам практично з усіма ядерними центрами таламуса здібна надавати регулюючий вплив на їх функціональну активність. ЕПІТАЛАМУС
Епіталамус (epithalamus) включає епіфіз (шишковидне тіло) і ряд ядерних скупчень нейронів. Епіфіз – залоза внутрішньої секреції, функція якої полягає у гальмуванні великої кількості інших ендокринних залоз (гіпофіза, щитовидної і прищитовидної залоз, статевих залоз, наднирників та інших). Епіфіз виробляє нейрогормон мелатонін, який має велике значення для підтримки імунного статуту організму. Гормони епіфіза також відіграють певну роль у регуляції сезонних ритмів життєдіяльності організму. Зовні епіфіз покритий тонкою сполучнотканинною капсулою, від якої до середини залози відходять перегородки, які утворюють строму залози і поділяють паренхіму на слабо виражені частки (мал. 42).
Мал. 42. Епіфіз людини: А-мале збільшення, Б-велике збільшення, 1-сполучнотканинна капсула, 2-сполучнотканинні перегородки, 3-паренхіма епіфіза, 4-мозковий пісок, 5-кровоносні судини, 6-гліоцити, 7-темні пінеалоцити, 8-світлі пінеалоцити (мал. В.Г. Єлисєєва).
Паренхіма епіфіза утворена клітинами двох типів – пінеальними і нейрогліальними. Пінеальні клітини (пінеалоцити) – залозисті, розміщуються в центрі часток. За формою вони багатокутні з пузирковидними ядрами. Серед пінеалоцитів розрізняють клітини із світлою і темною цитоплазмою. Темні клітини мають менші розміри і містять ацидофільні, а іноді базофільні гранули у цитоплазмі. Ці форми пінеалоцитів є не самостійними різновидами клітин, а свідчать про їх різний функціональний стан. Пінеалоцити мають відростки, які направляються до капілярів, і контактуючи з ними, розширюються. У цих розширеннях скупчуються осміофільні гранули, вакуолі і мітохондрії. По периферії часток скупчуються переважно гліальні клітини, які відрізняються бідною цитоплазмою і ущільненими ядрами.
МЕТАТАЛАМУС Метаталамус (metathalamus) або зазгір’я, розміщується у задньо-бічному відділі проміжного мозку, де під подушкою таламуса лежать два парних овальних утворення – більш велике присереднє і менше за розміром бічне колінчасті тіла. За допомогою ручок верхнього і нижнього горбиків, які складаються з білої речовини, присередні і бічні колінчасті тіла з’єднуються відповідно з нижніми і верхніми горбиками пластинки середнього мозку. Зверху колінчасті тіла покриті білою речовиною, усередині містять скупчення сірої речовини – ядра. Ядра присереднього колінчастого тіла (як і ядра нижнього горбика пластинки), є підкорковими центрами слуху, оскільки в них закінчуються аферентні волокна у складі бічної петлі, що беруть початок у ділянці моста (слуховий шлях) від ядер присінково - завиткового (VIII пара) нерву. Ядра бічного колінчастого тіла (разом з ядрами верхнього горбка пластинки середнього мозку) є підкорковими центрами зору. В них закінчується бічна частина волокон, які ідуть у складі зорового тракту (ІІ пара черепних нервів). Ядра колінчастих тіл формують також висхідні шляхи до центрів зорового і слухового аналізаторів у корі великих півкуль.
СУБТАЛАМУС
Субталамус (subthalamus) є підталамічною ділянкою, де закінчуються червоні ядра ніжок мозку і чорна речовина. Спереду і латерально від чорної речовини розміщується субталамічне ядро, або люїсове тіло. Це ядро відноситься до утворень екстрапірамідної системи мозку, яка забезпечує керування автоматизованими складнокоординованими рухами тіла. При локальних пошкодженнях субталамічного ядра виникають сильні “кидкові” рухи кінцівок (баллізм). ГІПОТАЛАМУС
Гіпоталамус (hypothalamus) або підзгір’я, є вентральним відділом проміжного мозку. До його складу входить комплекс утворень, який розташований під ІІІ шлуночком. Гіпоталамус спереду обмежується зоровим перехрестям (хіазмою), латерально – передньою частиною субталамуса, внутрішньою капсулою і зоровими трактами, що відходять від хіазми. Позаду гіпоталамус продовжується у покришці середнього мозку (мал. 43). У гіпоталамусі розрізняють велику переднью бічну частину і меншу задню частину. Передньо-бічна частина утворює дно ІІІ шлуночка мозку і включає сірий горб, лійку з гіпофізом, зорові тракти, перехрестя зорових нервів (хіазму), термінальну мозкову пластинку. Також сюди відносять власну підбугорну ділянку – скупчення ядер сірої речовини гіпоталамуса. Сірий горб – це тонка мозкова пластинка, яка розміщується між перехрестям зорових нервів спереду і сосочковими тілами позаду. У стінці сірого горба розташовані вегетативні ядра, які належать до емоціогенних зон мозку. Витончене продовження передньої стінки сірого горба догори і назад представляє собою пограничну пластинку, яка утворює передню стінку ІІІ шлуночка. Вентрально сірий горб утворює лійку, яка служить місцем приєднання гіпофіза. Підгорбова ділянка розташована під таламусом дещо вище сірого бугра і частково між його стінками. Зверху вона відокремлена від таламусів гіпоталамічною борозною. Підбугорна ділянка містить до 40 ядер, функціональна роль яких дуже важлива. У передньо-бічній частині гіпоталамуса розрізняють передню і середню групи гіпоталамічних ядер.
Мал. 43. Вентральна поверхня проміжного мозку: 1-зоровий нерв, 2-передня продірявлена речовина, 3-зорове перехрестя, 4-зоровий тракт, 5-міст, 6-міжніжкова ямка, 7-задня продірявлена речовина, 8-сосочкові тіла, 9-сірий горб, 10-лійка, 11-латеральна нюхова смужка, 12-нюховий трикутник, 13-закрутки лобової частки (мал. В.І. Козлова, Т.А. Цехмистренко).
До передньої групи відносять паравентрикулярне ядро, преоптичне ядро, супраоптичні і супрахіазматичні ядра. Паравентрикулярне ядро розміщується під епендимою уздовж стінки ІІІ шлуночка, преоптичне ядро – спереду ближче до латеральної стінки гіпоталамуса над сірим горбом, група супраоптичних і супрахіазматичних ядер – над хіазмою і зоровим трактом своєї сторони. У нейронах паравентрикулярного і супраоптичного ядер утворюється нейросекрет, який по їх аксонах переміщується у задній відділ гіпофіза (нейрогіпофіз), де виводиться в кров у вигляді нейрогормонів – вазопресина і окситоцина. Вазопресин (антидіуретичний гормон) стимулює зворотне всмоктування (резорбцію) води в ниркових канальцях. Пошкодження передніх ядер приводить до припинення виведення вазопресину, в результаті чого розвивається нецукровий діабет. Окситоцин стимулює скорочення гладенької м’язової тканини внутрішніх органів. У цілому від цих гормонів залежить водно-сольовий обмін в організмі. У преоптичному ядрі утворюється один із рилізінг-гормонів - люліберин, який стимулює виробку в аденогіпофізі лютеїнізуючого гормону, що контролює активність статевих залоз. Супрахіазматичні ядра приймають участь у регуляції циклічних змін фізіологічних процесів в організмі (біоритмів). Відомо, що стимуляція переднього відділу гіпоталамуса приводить до реакцій парасимпатичного типу (звуження зіниці, зниження частоти серцевих скорочень, розширення просвіту судин і падіння артеріального тиску, посилення перистальтики кишечнику). Руйнування цього відділу супроводжується також незворотнім підвищенням температури тіла.
Мал. 44. Схема гіпоталамо-гіпофізарної системи і регуляції тропними гормонами: 1-хіазма, 2-медіальна еміненція з первинною капілярною сіткою, 3-порожнина ІІІ шлуночка, 4-супраоптичне ядро, 5-преоптична зона гіпоталамуса, 6-паравентрикулярне ядро, 7-ядра середнього відділу гіпоталамуса, 8-клітини ядер середнього відділу гіпоталамуса, які виробляють рилізінг-фактори в первинну капілярну сітку медіальної еміненції, 9-адренергічні нейрони середнього гіпоталамусу, які дають початок низхідним еферентним нервовим шляхам (парааденогіпофізарна передача гіпоталамічних імпульсів до регулюємих ефекторів), 10-таламус, 11-лійка ІІІ шлуночка і гіпофізарна ніжка, 12-задня частка гіпофіза, 13-накопичувальне тільце Херринга (закінчення аксонів клітин ядер переднього гіпоталамуса), 14-середня частка гіпофіза, 15-гіпофізарна щілина, 16-передня частка гіпофіза із вторинною капілярною сіткою, 17-портальна (ворітна) вена, 18-туберальна частина аденогіпофіза. Аденогіпофізарні гормони та їх органи-мішені: СТГ - стимулює ріст організму в цілому та його окремих органів, АКТГ – стимулює пучкову і сітчасту зони кори надниркових залоз, ЛГ – стимулює овуляцію, утворення жовтого тіла яєчника і продукцію прогестерону, стимулює продукцію тестостерону в сім’яниках, ФСГ – активує ріст фолікулів і виробку ними естрогенів в яєчниках, стимулює сперматогенез у сім’яниках, ТТГ – активує продукцію і секрецію тироїдних гормонів щитовидної залози, ЛТГ – активує виробку молока в молочних залозах. ОКС – викликає скорочення матки і активує синтез молока у молочних залозах, АДГ – стимулює зворотну реабсорбцію води із первинної сечі в нирках і одночасно підвищує артеріальний тиск крові (мал. Б.В. Алешина).
До середньої групи гіпоталамічних ядер відносять дорсо-медіальне і вентро-медіальне ядра, ядро сірого горба і ядро лійки. Середня група ядер контролює водний, жировий і вуглеводний обмін, впливає на рівень цукру в крові, іонний баланс організму, проникність судин і клітинних мембран. В ядрах середньої групи локалізуються центри голоду і насичення. Наприклад, руйнування вентро-медіального ядра гіпоталамуса приводить до надлишкового споживання їжі (гіперфагії) і ожиріння, а пошкодження ядра сірого горба – до зниження апетиту і різкому схудненню (кахексії). Нейрони ядер цієї групи виробляють ряд рилізінг – гормонів (соматостатин, соматоліберин, люліберин, фоліберин, пролактоліберин, тироліберин та інші). Через гіпоталамо-гіпофізарну систему вони впливають на ростові процеси, швидкість фізичного розвитку і статевого дозрівання, формування вторинних статевих ознак, функції статевої системи, а також на обмін речовин (мал. 44). В цілому можна сказати, що ядра середньої групи мають відношення до регуляції харчової і статевої поведінки, а також до контролю функцій ендокринних залоз. Крім цього, в цій зоні гіпоталамуса знаходяться специфічні центри, які відіграють важливу роль у процесах формування мотивації і психоемоційних форм поведінки. Задня частина гіпоталамуса розташована між сірим горбом і задньою продірявленою речовиною і складається з правого і лівого сосочкових тіл. Усередині кожного з них під тонким шаром білої речовини знаходяться два сірих ядра, які відносяться до підкоркових нюхових центрів. До задньої групи ядер належить також заднє гіпоталамічне ядро. Разом з ядрами передньої групи заднє гіпоталамічне ядро приймає участь у терморегуляції, а також містить центри, що координують активність симпатичної частини автономної нервової системи. Стимуляція цього ядра приводить до реакцій симпатичного типу (розширення зіниці, підвищення частоти серцевих скорочень, і артеріального тиску, прискорення дихання і зменшення тонічних скорочень кишечнику). Зруйнування заднього відділу гіпоталамуса викликає сонливість, млявість і зниження температури тіла. Гіпоталамус має широку систему зв’язків з різними утвореннями головного мозку, що пояснює його участь у багатьох поведінкових реакціях. Система аферентних і еферентних волоконсклепіннязв’язує передній гіпоталамус і сосочкові тіла з гіпокампом великих півкуль, а сосочково-таламічний (пучок Вік д’Азіра) і сосочково - покривний шляхи зв’язують сосочкові тіла з таламусом і покришкою середнього мозку. Гіпофіз (hypophysis), нижній мозковий придаток або пітуїтарна залоза ( glandula pituitaria) – залоза внутрішньої секреції, яка розміщується в ямці турецького сідла клиновидної кістки черепа. У структурному і функціональному відношенні це один з найбільш складних органів внутрішньої секреції, про який кажуть, як про центральну залозу ендокринної системи. Така характеристика обумовлена тим, що нижній мозковий придаток займає центральне положення не тільки за своєю топографією в основі мозку, але і у функціональному відношенні є залозою, яка у певній мірі підпорядковує діяльність інших, так названих “периферичних ендокринних залоз”: щитовидної, статевих залоз, а також кори надниркових залоз. Гіпофіз приймає участь у регуляції основних біологічних процесів, характеризуючи життєвий цикл кожного індивідуума. Такі біологічні процеси як ріст і розмноження хребетних тварин у значній ступені залежать від нормальної діяльності гіпофіза. Крім цього, гіпофіз також приймає участь у регуляції всіх видів обміну речовин: водного, мінерального, вуглеводного, білкового і жирового. З усього різноманіття класифікацій будови гіпофіза, класичним є поділ його на аденогіпофіз і нейрогіпофіз. В аденогіпофізі, в свою чергу, розрізняють передню, середню (або проміжну) і туберальну частки. Аденогіпофіз представляє собою складну залозу, яка утворена тяжами епітеліальних клітин (аденоцитів), поділених розвинутою сіткою синусоїдних капілярів. Передня частка аденогіпофіза має трабекулярну будову. Трабекули, дуже розгалужуються, сплітаються у вузькопетлисту сітку. Проміжки між ними заповнені пухкою сполучною тканиною, по якій проходять численні синусоїдні капіляри. У кожній трабекулі можна розрізнити декілька різновидів залозистих клітин. Одні з них, що розташовані по периферії трабекул, більш великі за розмірами, містять секреторні гранули і інтенсивно фарбуються на гістологічних препаратах. Тому ці клітини іменують хромофільними (cellulae chromophilica). Другі клітини хромофобні (cellulae chromophobicae), займають середину трабекул, відрізняються від хромофільних клітин слабо забарвленою цитоплазмою. У зв’язку з кількісним переваженням у складі трабекул хромофобних клітин їх іноді називають головними. Хоча у функціональному відношенні, як показують дослідження вони мало активні. Хромофільні клітини поділяють на базофільні (cellulae basophilicae) і ацидофільні (cellulae acidophilicae). Базофільні клітини або базофіли виробляють глюкопротеїдні гормони і їх секреторні гранули на гістологічних препаратах фарбуються основними барвниками. Сумарний вміст базофілів коливається від 4 до 10% від загальної кількості аденоцитів передньої частки гіпофіза. Базофіли порівняно великі за розмірами. Серед них розрізняють дві основні різновидності: гонадотропи (cellulae gonadotrophica) - виробляють фолікулостимулюючий (фолікулотропін) і лютеїнізуючий гормони (лютропін), що впливають на функції статевих залоз та тиротропи (cellulae thyrotropica) - виробляють тиротропний гормон (тиротропін), який регулює функціональну активність щитовидної залози (мал. 45).
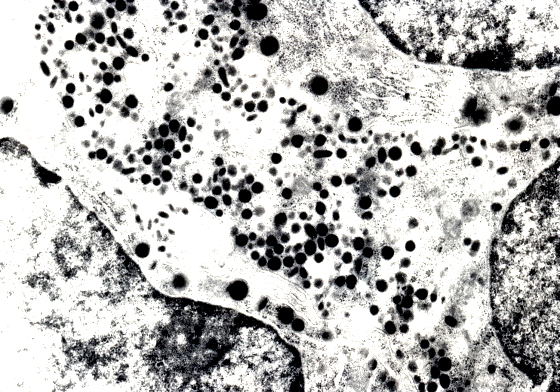
Мал. 45. Аденогіпофіз. Тиротроп (1) і фрагменти двох лактотропів (2, 3). Електронограма. Збільшення х 8.000.
Для ацидофільних клітин характерні великі щільні гранули, які фарбуються на гістологічних препаратах кислими барвниками. Ацидофіли складають 30-35%усіх аденоцитів, відрізняються дещо меншими розмірами порівняно базофільних клітин і не містять глюкопротеїдів. Ацидофільні клітини поділяють на дві різновидності: соматотропи (cellulae comatotropicae), які виробляють соматотропний гормон (соматотропін), що впливає на білковий обмін і ріст трубчастих кісток і лактотропи (cellulae mammotropicae) – виробляють лактотропний гормон, який стимулює розвиток тканин і лактацію. Крім того, в аденогіпофізі серед хромофобів виділяють також кортикотропні клітини (cellulae corticotropicae), які виробляють адренокортикотропний гормон (активізує діяльність кори надниркових залоз) і так званні фолікулярно-зірчасті клітини (мал. 46). Середня частка аденогіпофіза утворена декількома рядами залозистих клітин (від 5 до 15). Аденоцити середньої частки здібні виробляти білковий секрет, який накопичується між сусідніми клітинами, приводить до формування у середній частці фолікулоподібних порожнин. Іноді, вони заповнюються колоїдоподібною речовиною. Поряд з цими порожнинами, у тварин, які мають гіпофізарну щілину, колоїдоподібні маси можуть накопичуватись у ній. У середній частці аденогіпофіза виробляються меланотропний гормон (меланотропін), що впливає на пігментний обмін, а також ліпотропний гормон (посилює метаболізм жироподібних речовин).
Мал. 46. Електронномікроскопічна будова клітин аденогіпофіза: І-гонадотропна фолікулостимулююча клітина, ІІ-тиротропна клітина, ІІІ-соматотропна клітина, IV-лактотропна клітина, V-кортикотропна клітина, VI-клітина тироїдектомії, VII-клітина кастрації, VIII-фолікулярно-зірчаста клітина, 1-ядро, 2-комплекс Гольджі, 3-мітохондрії, 4-ендоплазматична сітка, 5-секреторні гранули, 6-вакуоль, 7-мікроворсинки, 8-порожнина фолікула, яка заповнена колоїдоподібною масою (мал. Б.В. Алешина).
Частину аденогіпофіза, що прилягає до гіпофізарної ніжки і стикається з нижньою поверхнею медіальної еміненції гіпоталамуса називають – туберальною. За структурою туберальна частина нагадує середню частку і вона утворена кубічними або призматичними епітеліальними клітинами з помірно базофільною цитоплазмою. Від туберальної частини, також як і від епітелію середньої частки, відходять трабекули, які складають паренхіму передньої частки гіпофіза, тому в деяких клітинах туберальної частини зустрічаються базофільні гранули, але не у великій кількості. Задня частка гіпофіза утворена нейроглією. Основним джерелом гліальних волокон задньої частки є епендимні клітини, які покривають порожнину лійки, залишки якої іноді зберігаються у середині задньої частки. Гліальні клітини цієї частки представлені переважно невеликими виростковими або веретеноподібними клітинами - пітуїцитами. Численні відростки цих клітин закінчуються на адвентиції судин і базальній мембрані капілярів. До задньої частки входять аксони нейросекреторних клітин супраоптичного і паравентрикулярного ядер переднього гіпоталамуса, які збираються у гіпоталамо-нейрогіпофізарні нервові пучки і проходять через медіальну еміненцію і гіпофізарну ніжку. У задній частці ці волокна закінчуються розширеними терміналями (тільця Херингу), що контактують з капілярами. Пітуїцити мають численні цитоплазматичні відростки, які проникають між нейросекреторними волокнами та їх розширеннями і тісно контактують з ними. Тіла пітуїцитів також тісно контактують з безліччю нейросекреторних розширень, утворюючи з ними синаптоподібні контакти. Часто, такі розширення глибоко проникають у цитоплазму пітуїцитів, а іноді оказуються усередині неї, поблизу ядра. Розміри, форма і внутрішня структура пітуїцитів значно варіює, що дало підставу виділити декілька типів цих клітин: астро, - оліго, - і мікропітуїцити.
КІНЦЕВИЙ МОЗОК
Кінцевий мозок (telencephalon), або великий мозок, представляє собою саму розвинену і у філогенетичному відношенні нову частину головного мозку, яка безпосередньо зв’язана з найбільш складними проявами психічної та інтелектуальної діяльності людини. Кінцевий мозок складається з двох півкуль великого мозку, які з’єднані між собою мозолистим тілом, передньою і задньою спайками і спайкою склепіння. Порожнини кінцевого мозку складають правий і лівий бічні шлуночки, кожний з яких знаходиться у відповідній півкулі. Медіальну стінку бічного шлуночка в ростральному відділі утворює прозора перегородка. Півкулі великого мозку зверху покрити корою мозку – шаром сірої речовини, який утворений нейронами понад п’ятдесяти різновидів. Під корою мозку у великих півкулях знаходиться біла речовина, що складається з мієлінових волокон, більша частина яких з’єднує кору великих півкуль з іншими відділами і центрами головного мозку. У товщі білої речовини півкуль знаходяться скупчення сірої речовини – базальні ядра. Шар білої речовини (внутрішня капсула) обмежує півкулі від таламусів проміжного мозку.
ПІВКУЛІ ВЕЛИКОГО МОЗКУ
Права і ліва півкулі мозку відокремлені одна від одної повздовжньою щілиною. У кожній півкулі розрізняють три поверхні – латеральну (бічну), медіальну (внутрішню) і нижню, і три полюса – лобовий, потиличний і висковий. Поверхня плащової частини кожної півкулі поділяється за допомогою щілин і борозен на частки і закрутки (мал. 47). Щілини, або первинні борозни, як правило, глибокі і належать до постійних утворень мозку. Вони з’являються на 5-му місяці внутрішньоутробного розвитку і поділяють півкулі на частки. Рельєф поверхні півкуль багато в чому визначається вторинними і третинними борознами. Їх формування відбувається від моменту народження до 7-8 років і підлягає індивідуальним змінам. Третинні борозни особливо відрізняються індивідуальним рисунком. Однак, у більшості людей рельєф півкуль, зокрема розташування глибоких постійних борозен, має схожий характер.
3 9
6 12
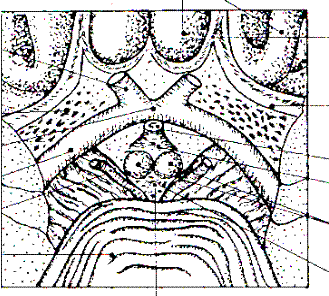
Мал. 47. Рельєф півкуль головного мозку (вигляд зверху): 1-повздовжня щілина мозку, 2-верхня лобова борозна, 3-нижня лобова борозна, 4-передцентральна борозна, 5-центральна борозна, 6-зацентральна борозна, 7-внутрішньо-тім’яна борозна, 8-верхня лобова закрутка, 9-середня лобова закрутка, 10-передцентральна закрутка, 11-зацентральна закрутка, 12-верхня тім’яна частка, 13-нижня тім’яна часточка (мал. В.І. Козлова, Т.А. Цехмистренко)..
Великі борозни і щілини поділяють кожну півкулю на 6 часток: лобову,тім’яну, потиличну, вискову, острівцеву і лімбічну. На латеральній поверхні півкулі розрізняють центральну (роландову) борозну і бічну (сільвієву) борозну. Попереду від центральної борозни розташована лобова частка, позаду – тім’яна. Знизу лобова частка півкулі відокремлюється завдяки бічній борозні від вискової частки. У верхній частині бічної поверхні півкулі тім’яна частка відокремлена від потиличної частки тім’яно-потиличною борозною. Передньо - нижньою межею потиличної частки є умовна лінія, яка проходить від верхнього кінця тім’яно-потиличної борозни донизу до нижнього краю півкулі. У глибині бічної борозни знаходиться острівцева частка (або островець). Ця частка покрита частинами лобової, тім’яної і вискової часток. На медіальній поверхні півкулі поряд з мозолистим тілом розташована його лімбічна частка, яка відокремлена від інших часток поясною закруткою.
Мал. 48. Медіальна поверхня правої півкулі великого мозку, мозкового стовбуру і мозочка. Водопровід середнього мозку, IV шлуночок. 1-підмозолисте тіло, 2-задня нюхова борозна. 3-нюхова площадка, 4-прозора перегородка. 5,11-поясна борозна, 6-верхня лобова закрутка, 7-коліно мозолистого тіла, 8-поясна закрутка, 9-борозна мозолистого тіла. 10-стовбур мозолистого тіла, 12-центральна борозна, 13-прицентральна часточка, 14-передклин, 15-тім’яно-потилична борозна, 16-підтім’яна б
|
|||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2017-01-26; просмотров: 529; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.142.55.138 (0.017 с.) |





 1 12
1 12 2 11
2 11 3
3 4 9
4 9 5
5

 12
12 3
3






