
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Исторические этапы развития генетики как науки.Содержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
ОТВЕТЫ К ЭКЗАМЕНУ ПО БИОЛОГИИ. 2011-2012 УЧ.ГОД. Раздел №2 Генетика. Исторические этапы развития генетики как науки. Первый этап ознаменовался открытием Г. Менделем (1865) дискретности (делимости) наследственных факторов и разработкой гибридологического метода, изучения наследственности, т. е. правил скрещивания организмов и учета признаков у их потомства. Второй этап характеризуется переходом к изучению явлений наследственности на клеточном уровне (питогенетика). Т. Бовери (1902—1907), У. Сэттон и Э. Вильсон (1902—1907) установили взаимосвязь между менделевскими законами наследования и распределением хромосом в процессе клеточного деления (митоз) и созревания половых клеток (мейоз). В этом периоде было установлено, что гены расположены в хромосомах в линейном порядке, образуя группы сцепления. Число групп сцепления генов соответствует числу пар гомологичных хромосом, и гены одной группы сцепления могут перекомбинироваться в процессе мейоза благодаря явлению кроссинговера. Также были установлены закономерности наследования признаков, сцепленных с полом. Третий этап в развитии генетики отражает достижения молекулярной биологии и связан с использованием методов и принципов точных наук — физики, химии, математики, биофизики и др.—в изучении явлений жизни на уровне молекул. Объектами генетических исследований стали грибы, бактерии, вирусы. На этом этапе были изучены взаимоотношения между генами и ферментами и сформулирована теория “один ген — один фермент” (Дж. Бидл и Э. Татум, 1940): каждый ген контролирует синтез одного фермента; фермент в свою очередь контролирует одну реакцию из целого ряда биохимических превращений, лежащих в основе проявления внешнего или внутреннего признака организма. Эта теория сыграла важную роль в выяснении физической природы гена как элемента наследственной информации. В последнее десятилетие возникло новое направление в молекулярной генетике —генная инженерия — система приемов, позволяющих биологу конструировать искусственные генетические системы. Генная инженерия основывается на универсальности генетического кода. Таким образом, третий, современный этап развития генетики открыл огромные перспективы направленного вмешательства в явления наследственности и селекции растительных и животных организмов, выявил важную роль генетики в медицине, в частности, в изучении закономерностей наследственных болезней и физических аномалий человека.
2.2 История изучения ДНК ДНК была открыта Иоганном Фридрихом Мишером в 1869 году. Из остатков клеток, содержащихся в гное, он выделил вещество, в состав которого входят азот и фосфор. Вначале новое вещество получило название нуклеин, а позже, когда Мишер определил, что это вещество обладает кислотными свойствами, оно получило название нуклеиновая кислота. Биологическая функция новооткрытого вещества была неясна, и долгое время ДНК считалась запасником фосфора в организме. Даже в начале XX века многие биологи считали, что ДНК не имеет никакого отношения к передаче информации, поскольку строение молекулы, по их мнению, было слишком однообразным и не могло содержать закодированную информацию. Постепенно было доказано, что именно ДНК является носителем генетической информации. Одно из первых решающих доказательств принесли эксперименты О. Эвери, Колина Мак-Леода и Маклин Мак-Карти (1944 г.) по трансформации бактерий. Им удалось показать, что за так называемую трансформацию (приобретение болезнетворных свойств безвредной культурой в результате добавления в неё мёртвых болезнетворных бактерий) отвечают выделенные из пневмококков ДНК. Вплоть до 50-х годов XX века точное строение ДНК, как и способ передачи наследственной информации, оставалось неизвестным. Хотя и было доподлинно известно, что ДНК состоит из нескольких цепочек, состоящих из нуклеотидов, никто не знал точно, сколько этих цепочек и как они соединены. Химическая организация гена Исследования, направленные на выяснение химической природы наследственного материала, доказали, что материальным субстратом наследственности и изменчивости являются нуклеиновые кислоты, которые были обнаружены Ф. Мишером (1868) в ядрах клеток гноя. Нуклеиновые кислоты являются макромолекулами, т.е. отличаются большой молекулярной массой. Это полимеры, состоящие из мономеров - нуклеотидов, включающих три компонента: сахар (пентозу), фосфат и азотистое основание (пурин или пиримидин). К первому атому углерода в молекуле пентозы С-1 присоединяется азотистое основание (аденин, гуанин, цитозин, тимин или урацил), а к пятому атому углерода С-5 с помощью эфирной связи - фосфат; у третьего атома углерода С-3 всегда имеется гидроксильная группа - ОН. Соединение нуклеотидов в макромолекулу нуклеиновой кислоты происходит путем взаимодействия фосфата одного нуклеотида с гидроксилом другого так, что между ними устанавливается фосфодиэфирная связь. В результате образуется полинуклеотидная цепь. Остов цепи состоит из чередующихся молекул фосфата и сахара. К молекулам пентозы в положении С-1 присоединено одно из перечисленных выше азотистых оснований. Сборка полинуклеотидной цепи осуществляется при участии фермента полимеразы, который обеспечивает присоединение фосфатной группы следующего нуклеотида к гидроксильной группе, стоящей в положении 3, предыдущего нуклеотида. Благодаря отмеченной специфике действия названного фермента наращивание полинуклеотидной цепи происходит только на одном конце: там, где находится свободный гидроксил в положении 3. Начало цепи всегда несет фосфатную группу в положении 5. Это позволяет выделить в ней 5 и 3 -концы. Генетический код Белки практически всех живых организмов построены из аминокислот всего 20 видов. Эти аминокислоты называют каноническими. Каждый белок представляет собой цепочку или несколько цепочек аминокислот, соединённых в строго определённой последовательности. Эта последовательность определяет строение белка, а следовательно все его биологические свойства. Реализация генетической информации в живых клетках (то есть синтез белка, кодируемого геном) осуществляется при помощи двух матричных процессов: транскрипции (то есть синтеза мРНК на матрице ДНК) и трансляции генетического кода в аминокислотную последовательность (синтез полипептидной цепи на мРНК). Для кодирования 20 аминокислот, а также сигнала «стоп», означающего конец белковой последовательности, достаточно трёх последовательных нуклеотидов. Набор из трёх нуклеотидов называется триплетом. Принятые сокращения, соответствующие аминокислотам и кодонам, изображены на рисунке. Свойства 1. Триплетность — значащей единицей кода является сочетание трёх нуклеотидов (триплет, или кодон). 2. Непрерывность — между триплетами нет знаков препинания, то есть информация считывается непрерывно. 3. Неперекрываемость — один и тот же нуклеотид не может входить одновременно в состав двух или более триплетов (не соблюдается для некоторых перекрывающихся генов вирусов, митохондрий и бактерий, которые кодируют несколько белков, считывающихся со сдвигом рамки). 4. Однозначность (специфичность) — определённый кодон соответствует только одной аминокислоте (однако, кодон UGA у Euplotes crassus кодирует две аминокислоты — цистеин и селеноцистеин)[1] 5. Вырожденность (избыточность) — одной и той же аминокислоте может соответствовать несколько кодонов. 6. Универсальность — генетический код работает одинаково в организмах разного уровня сложности — от вирусов до человека (на этом основаны методы генной инженерии). 7. Помехоустойчивость — мутации замен нуклеотидов, не приводящие к смене класса кодируемой аминокислоты, называют консервативными; мутации замен нуклеотидов, приводящие к смене класса кодируемой аминокислоты, называют радикальными. Механизм репарации в ДНК. Система защиты клетки включает различные типы репарации поврежденной молекулы ДНК. Этот процесс может быть одноэтапным и многоэтапным, происходить как на свету, nак и в темноте. Например, при эксцизионной репарации, специальный фермент делает надрез возле поврежденного участка, а затем этот участок удаляется. На месте образовавшейся бреши происходит репаративный синтез ДНК по матрице неповрежденной цепи. Ферменты репликации в редких случаях ошибочно вставляют вдочернюю цепь не комплементарное основание. Ошибки репликации исправляют специальные ферменты с корректирующей функцией; они находят и удаляют некомплементарное основание. Затем происходит замена на основание, соответствующее правилу комплементарноста (А- Т, G - С).
Анализирующее скрещивание. По фенотипу особи далеко не всегда можно определить ее генотип. У самоопыляющихся растений генотип можно определить в следующем поколении. Для видов, использующих другие системы полового размножения, применяют так называемое анализирующее скрещивание. Скрещивание гибридной особи с особью, гомозиготной по рецессивным аллелям, называется анализирующим. При анализирующем скрещивании особь, генотип которой следует определить, скрещивают с особями, гомозиготными по рецессивному гену, т.е. имеющими генотип аа. Анализирующее скрещивание – один из основных методов, позволяющих установить генотип особи, по этой причине оно широко используется в генетике и селекции. Неполное доминирование. Далеко не всегда гетерозиготные организмы по фенотипу точно соответствуют родителю, гомозиготному по доминантному гену. Случаи, когда гетерозиготные потомки имеют промежуточный фенотип, называют неполным доминированием. Неполное доминирование ни в коей степени не отменяет закон расщепления, но при неполном доминировании в потомстве гибрида (F2) расщепление по фенотипу и генотипу совпадает, поскольку гетерозиготные особи (Аа) отличаются по внешнему виду от гомозигот (АА). Неполное доминирование или, как еще говорят, промежуточное проявление признака широко распространено в природе.
Эпистаз.
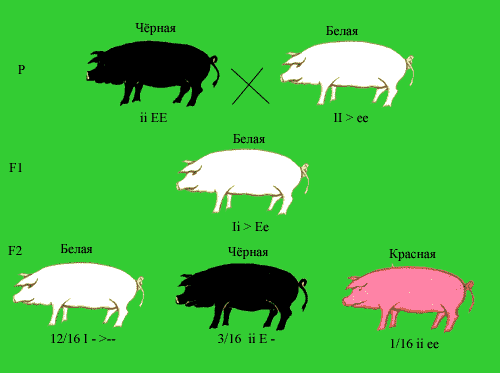
Эпистатическое взаимодействие неаллельных генов в определенном смысле противоположно комплементарному действию генов. Сущность эпистаза сводится к подавлению проявления генов одной аллельной пары генами другой. Гены, подавляющие действие других неаллельных генов, называются супрессорами или подавителями. Они могут быть как доминантными, так и рецессивными, например А - В- или bbA -. Наследование окраски у свиней демонстрирует доминантный Эпистаз. При скрещивании черных и белых свиней из разных пород в F1 появляются белые потомки. Их скрещивание между собой приводит к появлению белых (12/16), черных (3/16) и красных (1/16) поросят. Все белые поросята имеют минимум один доминантный генподавитель I. Черные поросята гомозиготны по рецессивному аллелю i, не препятствующему формированию окраски, и несут доминантный аллель Е, детерминирующий образование черного пигмента. Красные поросята (eeii) лишены доминантного геноподавителя I и доминантного гена, определяющего черную окраску. Полимерия. В некоторых случаях установлено, что проявление конкретного признака зависит от количества доминантных генов, вносящих вклад в его развитие. Например, при скрещивании краснозерных пшениц с белозерными было установлено, что растения с генотипом А1А1А2А2 имеют красные зерна, растения а1а1а2а2 – белые зерна, растения с тремя доминантными генами – красноватую окраску, а растения с двумя и одним геном – более бледную окраску. Таким образом, накопление определенных аллелей в генотипе может вести к изменению выраженности признаков.
2.25Плейотропное действие генов - это зависимость нескольких признаков от одного гена, то есть множественное действие одного гена. В дрозофилы ген белого цвета глаз одновременно влияет на цвет тела, длины, крыльев, строение полового аппарата, снижает плодовитость, уменьшает продолжительность жизни. У человека известна наследственная болезнь - арахнодактилия ("паучьи пальцы"-очень тонкие и длинные пальцы), или болезнь Марфана. Ген, отвечающий за эту болезнь, вызывает нарушение развития соединительной ткани и одновременно влияет на развитие нескольких признаков: нарушение строения хрусталика глаза, аномалии в сердечно-сосудистой системе.
Множественные аллели Три или большее число генов, которые могут находиться в одном и том же локусе, т. е. занимать одно и то же положение в гомологичных хромосомах, называют серией множественных аллелей. Все они обусловливают различные фенотипы. Каждая особь в популяции может иметь любые два из таких аллелей, но никак не больше, а каждая гамета, разумеется, содержит только один из них. Однако в популяции в целом соответствующий локус может быть представлен тремя или большим числом аллелей. По типу множественных аллелей наследуются группы крови О, А, В и АВ у человека. Ген IA содержит код для синтеза в эритроцитах специфического белка — агглютиногена А; ген IB вызывает синтез другого белка — агглютиногена В, ген i не продуцирует никакого агглютиногена. Ген i рецессивен по отношению к двум другим, но ни ген IA, ни ген IB не доминируют друг над другом. (Символы IA, IB и i указывают на то, что все эти три гена — аллели одного и того же локуса.) Таким образом, генотипы IA IA и IA i обусловливают группу крови А, генотипы IB IB и IB i — группу В, а генотип ii — группу О. Если же у человека имеются оба нерецессивных гена IA и IB, то у него образуются оба агглютиногена и он имеет группу крови АВ. Поскольку группы крови обусловлены генетически и не изменяются в течение всей жизни, их определение может помочь установить истину в случаях спорного отцовства. Однако на основании группы крови нельзя доказать, что данный человек действительно является отцом данного ребенка; можно лишь установить, что он мог бы быть его отцом, или же, исключить такую возможность. Подумайте: может ли мужчина с группой крови АВ быть отцом ребенка с группой О? Может ли ребенок с группой АВ быть сыном мужчины с группой О? Может ли ребенок с группой В иметь мать с группой А и отца с группой А или О? Наряду с системой АВО существует еще с десяток других систем групп крови, в том числе система MN и серия аллелей Rh; они определяются другими генами и наследуются независимо от групп АВО. Определение всех этих групп дозволяет иногда выяснять родственные отношения, которые нельзя было бы достоверно установить на основании одних лишь групп системы АВО.
Бомбейский феномен Бомбейский феномен заключается в том, что у ребенка определяется группа крови, которой по правилам у него быть не может - т.е. у ребенка выявляется антиген, которого нет ни у одного из родителей. Например, у родителей 00 и 00 (1-ая группа крови у обоих) вдруг рождается ребенок В0 (3-ья группа крови). Или у родителей с 00(1-ая гр.кр.) и B0/BB (3-ья гр.кр.) рождается ребенок с A0(2-ая гр.кр.) или AB (4-ая гр.кр.) Наследование резус-фактора
Резус-фактор - это липопротеид, расположенный на мембранах эритроцитов у 85% людей (они считаются резус-положительными). В случае его отсутствия говорят о резус-отрицательной крови. Эти показатели обозначаются латинскими буквами Rh со знаком «плюс» или «минус» соответственно. Для исследования резуса, как правило, рассматривают одну пару генов.
Родители: Dd x Dd. Дети: DD, Dd, dd. Гетерозиготность возникает как результат рождения резус-конфликтного ребенка у резус-отрицательной матери или может сохраняться в генах на протяжении многих поколений. Наследование группы крови системы Rh ребенком в зависимости от групп крови его родителей:
2.33 Моногибридное скрещивание Моногибридное скрещивание — скрещивание форм, отличающихся друг от друга по одной паре альтернативных признаков. При этом скрещиваемые предки являются гетерозиготными по положению хромосомы в аллели.
Рисунок 1: Шаблон, показывающий наследование доминантных (красного) и рецессивного (белый) фенотипов, когда каждый родитель (1) гомозиготен для доминантного либо рецессивного признака. Все члены I поколения гетерозиготны и имеют один и тот же общий для всех фенотип (2), в то время как поколение II показывает соотношение 3:1 доминантного к рецессивному фенотипам (3).
Моногибридное наследование представляет собой пример наследования единственного признака (гена), различные формы которого называют аллелями. Например, при моногибридном скрещивании между двумя чистыми линиями растений, гомозиготных по соответствующим признакам -- одного с жёлтыми семенами (доминантный признак), а другого с зелёными семенами (рецессивный признак), можно ожидать, что первое поколение будет только с жёлтыми семенами, потому что аллель жёлтых семян доминирует над аллелью зелёных. При моногибридном скрещивании сравнивают только один характерный признак.
Метод дерматоглифики Сущность метода состоит в анализе кожных узоров (рисунков) на ладонях и стопах. Метод наиболее информативен при хромосомных синдромах, когда выявляются дистальный осевой трирадиус, избыток дуг на пальцах, отсутствие дистальной межфаланговой складки, радиальные петли на I, IV и V пальцах, четырехпальцевая (обезьянья) складка (при болезни Дауна на коже ладоней у ребенка отмечается в 40-60% случаев). Аномалии числа хромосом Генетическая рекомбинация. Примером является мейотическая рекомбинация (кроссинговер) у эукариот, которая происходит в клетках после репликации, в профазе первого мейотического деления. Во время лептотены хромосомы конденсируются и становятся видимыми, В каждой из них после репликации дуплексная ДН К представлена двумя сестринскими хроматидами. Под электронным микроскопом видно, что на стадии лептотены пара сестринских хроматид каждой хромосомы формирует единый осевой элемент. Установлено, что у млекопитающих он состоит из белков SCP2 и SCP3 (от англ. synaptonemal complex proteins). На следующей стадии, зиготене, гомологичные хромосомы начинают соприкасаться друг с другом (конъюгировать) на отдельных, пока еще коротких участках. Одновременно осевые элементы гомологичных хромосом начинают соединяться попарно с помощью белка SCP1, который протягивается поперек между ними в виде субмикроскопических волокон (филамент). По завершении конъюгации, на стадии пахитены гомологичные хромосомы оказываются объединенными в биваленты по всей длине за счет специфической структуры, состоящей из двух продольных белковых тяжей. Это - так называемые латеральные элементы, в состав которых входят осевые элементы с прикрепленными к ним, петлеобразно уложенными фибриллами хроматина сестринских хроматид. Латеральные элементы соединены между собой поперечными белковыми волокнами, которые в совокупности формируют третью продольную структуру - центральный элемент. Из двух латеральных и одного центрального элемента образуется электроноплотная трехполосная структура, так называемый, синаптонемный комплекс, в котором гомологичные хромосомы прилегают к латеральным элементам с двух сторон, и этот контакт происходит «точечно» (в местах прикрепления петель к синаптонемному комплексу по всей era длине). Функциональное значение этой структуры, напоминающей застежку «молнию», состоит в том, что, с одной стороны, она не дает конъюгирующим хромосомам необратимо слипнуться, а с другой стороны — закрепляет их в строго гомологичном относительно локализованных на них генах взаиморасположении. В зависимости от размера генома у разных видов могут варьировать размеры синаптонсмного комплекса: общая ширина его трехполосной ленты составляет от 76 до 240 нм, а длина соответствует также видоспецифичной длине бивалентов в профазе 1 мейоза. На стадии диплотены гомологичные хромосомы бивалентов начинают расходиться, но обнаруживается, что несестринские хроматиды в биваленте остаются сцепленными в некоторых точках, образуя фигуру, получившую название хиазмы. На стадии диакинеза хромосомы конденсируются путем спирализации, а хиазмы вследствие отталкивания гомологов начинают сдвигаться к краям хромосом. В этот момент все четыре хроматиды становятся видимыми. Это — прямые наблюдения, и они позволяют сделать некоторые предположения о процессе рекомбинации. Профаза первого деления мейоза — единственный момент, когда гомологичные хромосомы образуют комплекс друг с другом, что, является условием, необходимым для осуществления рекомбинации. Можно полагать, что именно в обеспечении рекомбинации и состоит суть синапса—образование синаптонемногокомгаткса, временной структуры, которая формируется на стадии зиготены и разрушается в диплотене. Согласно мнению авторов приведенной выше гипотетической схемы, синаптонемный комплекс «...необходим для организации хроматина ввиде серии латеральных петель, основания которых собраны в линейную последовательность на поверхности его латеральных элементов и доступны для узнавания гомологичных локусов и кроссинговера». Существует очень правдоподобная, но до настоящего времени не всеми исследователями разделяемая гипотеза, что хиазмы представляют собой места прохождения рекомбинаций- ведь количество тех и других примерно совпадает. Это позволяет локализовать время происхождения процесса рекомбинаций, и считать, что даже на стадии диакинеза в местах рекомбинации хромосомы все еще остаются связанными посредством нитей ДНК. 2.50. Генотипическая изменчивость. Хромосомные аберрации, примеры Наследственная (генотипическая) изменчивость составляет часть общей (фенотипической) изменчивости, которая определяется генетическими различиями между особями или группами особей. Ведущую роль в процессах генотипической И. играет изменение генотипа в результате изменения репродуцирующихся структурных элементов ядра (ДНК хромосом) — мутации. Генотипы являются не чем иным, как комбинациями генов. Поэтому биологическое значение полового размножения заключается именно в том, что образование разнообразных генотипов при таком размножении значительно ускоряется. Наряду с мутациями источником генотипической И. являются рекомбинации генов (так называемая рекомбинационная И.). На степень генотипической И. в популяции влияет также приток генов из других популяций. Изменения генотипа приводят к изменению характера развития того или иного признака в данных условиях окружающей среды, изменению наследственной нормы реакции и появлению различных отклонений (наследственные аномалии развития). Сходство врожденных пороков развития, возникающих в результате действия повреждающих факторов окружающей среды, и наследственных аномалий объясняется тем, что при мутациях и под влиянием тератогенов в клетках формирующихся тканей и органов нарушаются одни и те же звенья биохимических процессов. Хромосомные аберрации (хромосомные мутации, хромосомные перестройки) — тип мутаций, которые изменяют структуру хромосом. Классифицируют делеции (утрата участка хромосомы),инверсии (изменение порядка генов участка хромосомы на обратный), дупликации (повторение участка хромосомы), транслокации (перенос участка хромосомы на другую), а также дицентрические и кольцевые хромосомы. Известны такжеизохромосомы, несущие два одинаковых плеча. Если перестройка изменяет структуру одной хромосомы, то такую перестройку называют внутрихромосомной (инверсии, делеции, дупликации, кольцевые хромосомы), если же двух разных, то межхромосомной (дупликации, транслокации, дицентрические хромосомы). Хромосомные перестройки подразделяют также на сбалансированные и несбалансированные. Сбалансированные перестройки (инверсии, реципрокные транслокации) не приводят к потере или добавлению генетического материала при формировании, поэтому их носители, как правило, фенотипически нормальны. Несбалансированные перестройки (делеции и дупликации) меняют дозовое соотношение генов, и, как правило, их носительство сопряжено с клиническими отклонениями от нормы. Хромосомные перестройки играют определенную роль в эволюционном процессе и видообразовании, в нарушении фертильности, в онкологических и врождённых наследственных заболеваниях человека. Хромосомные перестройки были открыты у дрозофил при помощи генетического анализа. В некоторых скрещиваниях соотношение числа потомков в разных классах сильно отличалось от ожидаемого, и это объяснили наличием перестроек в хромосомах родителей. Делеции, дупликации и транслокации обнаружил К. Бриджес в 1916, 1919 и 1923 годах, соответственно. Первую инверсию описал А. Стёртевант в 1921 году, сравнивая порядок генов в хромосоме 3 у D.melanogaster и D.simulans. Первые наблюдения хромосомных перестроек были сделаны на политенных хромосомах слюнных желез. Лишь спустя некоторое время существование перестроек было доказано цитологически на митотических хромосомах. Однако проще всего перестройки можно увидеть в политенных хромосомах у гетерозиготных особей, благодаря образованию петель и крестообразных структур. Также перестройки можно увидеть в профазе мейоза при образовании синаптонемных комплексов, где, благодаря синапсису гомологичных хромосом, также образуются петли и крестообразные структуры. Классификация Делеции
Некоторые типы хромосомных перестроек Различают терминальные (утрата концевого участка хромосомы) и интеркалярные (утрата участка на внутреннем участке хромосомы) делеции. Если после образования делеции хромосома сохранила центромеру, она аналогично другим хромосомам передается при митозе, участки же без центромеры, как правило, утрачиваются. При конъюгации гомологичных хромосом во время мейоза у нормальной хромосомы на месте, соответствующем интеркалярной делеции у дефектной хромосомы, образуется делеционная петля, которая компенсирует отсутствие делетированного участка. Врождённые делеции у человека редко захватывает протяженные участки хромосом, обычно такие аберрации приводят к гибели эмбриона на ранних этапах развития. Самым хорошоизученным заболеванием, обусловленным достаточно крупной делецией, является синдром кошачьего крика, описанный в 1963 годуЖеромом Леженом. В его основе лежит делеция участка короткого плеча 5 хромосомы. Для больных характерен ряд отклонений от нормы: нарушение функцийсердечно-сосудистой, пищеварительной систем, недоразвитие гортани (с характерным криком, напоминающим кошачье мяуканье), общее отставание развития,умственная отсталость, лунообразное лицо с широко расставленными глазами. Синдром встречается у 1 новорожденного из 50000. Современные методы выявления хромосомных нарушений, прежде всего флуоресцентная гибридизация in situ, позволили установить связь между микроделециями хромосом и рядом врождённых синдромов. Микроделециями, в частности, обусловлены давно описанные синдром Прадера-Вилли исиндром Вильямса. Дупликации Дупликации представляют собой класс перестроек, который объединяет как внутри-, так и межхромосомные перестройки. Вообще, любая дупликация — это появление дополнительной копии участка хромосомы, которая может располагаться сразу за тем районом, который дуплицирован, тогда это тандемная дупликация, либо в новом месте или в другой хромосоме. Новая копия может образовать отдельную маленькую хромосому со своими собственными теломерами и центромерой, тогда это свободная дупликация[1]:2. Тандемные дупликации появляются в половых клетках при мейозе в результате неравного кроссинговера (в этом случае второй гомолог несет делецию) или в соматических клетках в результате неаллельной гомологичной рекомбинации при репарации двунитевого разрыва ДНК. В процессе кроссинговера у гетерозиготы при конъюгации хромосомы с тандемной дупликацией и нормальной хромосомы, как и при делеции, формируется компенсационная петля. Практически у всех организмов в норме наблюдается множественность генов, кодирующих рРНК (рибосомальную РНК). Это явление назвали избыточностью генов. Так у E. coli на рДНК (ДНК, кодирующее рРНК) приходится 0,4 % всего генома, что соответствует 5-10 копиям рибосомальных генов. Другой пример дупликации — мутация Bar у Drosophila, обнаруженная в 20-х годах XX века Т. Морганом и А. Стёртевантом. Мутация обусловлена дупликацией локуса 57.0 X-хромосомы. У нормальных самок (B+/B+) глаз имеет 800 фасеток, у гетерозиготных самок (B+/B) глаз имеет 350 фасеток, у гомозиготпо мутации (B/B) — всего 70 фасеток. Обнаружены также самки с трижды повторенным геном — double Bar (BD/B+). В 1970 году Сусумо Оно в монографии «Эволюция путем дупликации генов» разработал гипотезу об эволюционной роли дупликаций, поставляющих новые гены, не затрагивая при этом функций исходных генов. В пользу этой идеи говорит близость ряда генов по нуклеотидному составу, кодирующих разные продукты. Это трипсин и химотрипсин, гемоглобин и миоглобин и ряд других белков. Инверсии Инверсией называют поворот участка хромосомы на 180°. Различают парацентрические (инвертированный фрагмент лежит по одну сторону от центромеры) и перицентрические (инвертированный фрагмент лежит по разные стороны от центромеры) инверсии. При инверсиях не происходит потери генетического материала, поэтому инверсии, как правило, не влияют на фенотип носителя. Однако, если у гетерозигот по инверсиям (то есть у организма, н
|
|||||||||||||||
|
|
Последнее изменение этой страницы: 2016-08-16; просмотров: 1314; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.137.170.76 (0.024 с.) |


 Рождение ребенка с отрицательным резус-фактором в семье с резус- положительными родителями в лучшем случае вызывает глубокое недоумение, в худшем – недоверие. Упреки и сомнения в верности супруги. Как ни странно, ничего исключительного в этой ситуации нет. Существует простое объяснение такой щекотливой проблемы.
Рождение ребенка с отрицательным резус-фактором в семье с резус- положительными родителями в лучшем случае вызывает глубокое недоумение, в худшем – недоверие. Упреки и сомнения в верности супруги. Как ни странно, ничего исключительного в этой ситуации нет. Существует простое объяснение такой щекотливой проблемы.





