Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Отряд Двукрылые(по-латински). Характеристика. Основные семейства. Москиты, характеристика. Медицинское значение.Содержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте Отряд Двукрылые - Diptera. У представителей этого отряда пара крыльев, ротовые аппараты их имеют форму хоботка, предназначенного для высасывания жидкой пищи из растительных или животных тканей либо для слизывания её с поверхности. Развитие с полным метаморфозом. Наибольшее значение представители этого отряда имеют как временные кровососущие паразиты и переносчики возбудителей трансмиссивных заболеваний. Массовые виды этого отряда, размножаясь в летний период в тундре и тайге в огромном количестве, делают жизнь человека почти невыносимой. Разные виды кровососущих двукрылых объединяются общим понятием гнус. Самцы большинства кровососущих двукрылых питаются соками растений, а самки – преимущественно или исключительно кровью животных и человека. В большинстве случаев откладка яиц самками наступает только после кровососания. Москиты более мелкие насекомые длиной 1,5-3,5 мм, имеющие тонкий хободок, сильно выступающий в виде горбика грудной отдел тела и обильное опущение тела и крыльев мелкими щетинками. Встречаются в тропических и субтропических зонах на всех континентах. Они известны как переносчики разных видов лейшманий, вирусов лихорадки паппатачи и возбудителей ряда других трансмиссивных заболеваний. Вирус лихорадкипаппатачи передается в поколениях инвазированных москитов трансовариально. Москиты не способны к большим перелетам, поэтому природные очаги заболеваний, вызываемых перечисленными паразитами, обычно имеют небольшие размеры.
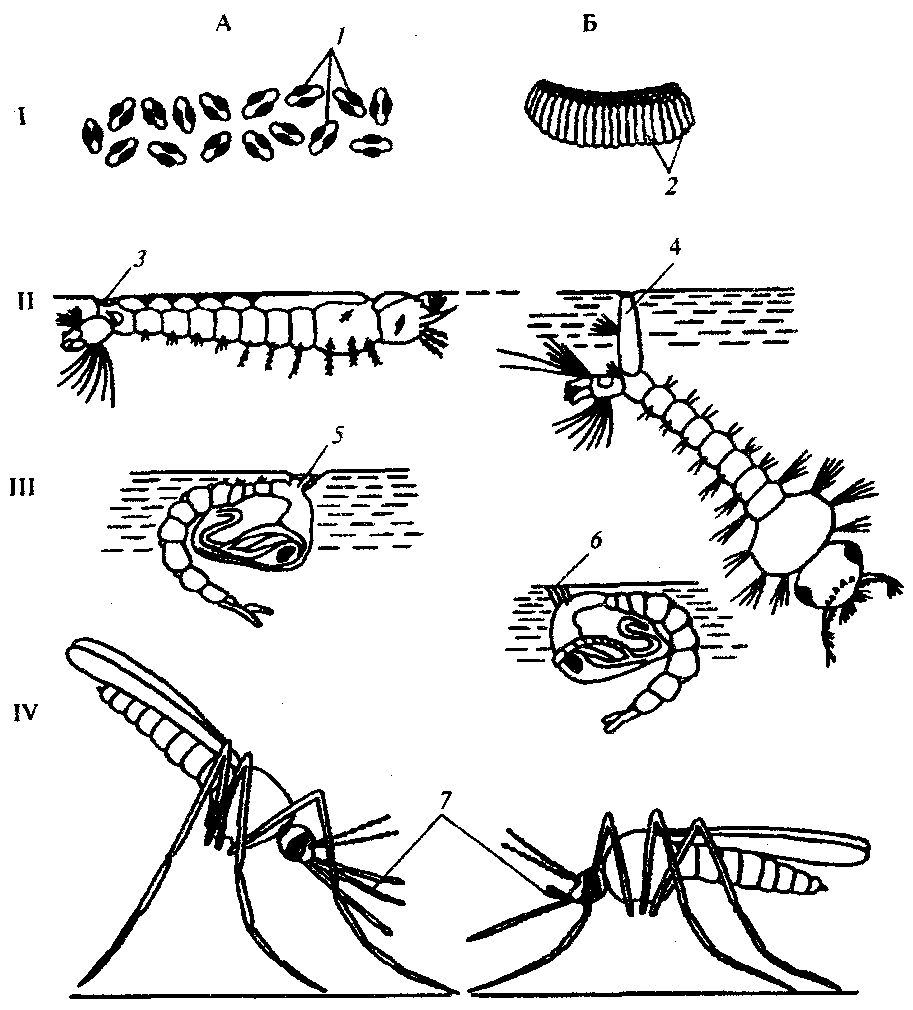
3.52: Комары (сем. Culicidae). Имеют тонкое стройное тело, длинные ноги и небольшую головку с ротовым аппаратом в виде длинного хоботка. Распространены повсеместно. Они сохраняют в организме и обладают способностью передавать животным и человеку возбудителей более 50 вирусных, бактериальных и паразитарных заболеваний. После оплодотворения самки активно ищут хозяина-прокормителя и способны обнаруживать его на расстоянии до 3 км с помощью обоняния, а затем и зрения. Больных с высокой температурой комары кусают более охотно. Слабое локальное освещение в вечерние и предутренние часы стимулирует двигательную активность комаров и поиск ими хозяев. В связи с этим комары в темное время суток часто влетают в освещенные окна и двери жилых и хозяйственных построек. В дневные часы суток при высокой температуре и низкой влажности воздуха комары обычно находятся в укрытиях и не питаются. Указанные особенности поведения комаров отражаются на адаптациях к паразитизму возбудителей заболеваний, передающихся комарами: при большинстве заболеваний этой группы у больных повышается температура в вечерние и ночные часы суток, в это же время в крови обнаруживается максимальная концентрация инвазионных для комаров стадий возбудителей. Комары откладывают яйца в воду или на влажную почву около воды. Личинки и куколки ведут водный образ жизни, а дышат атмосферным воздухом с помощью трахей. Личинки питаются взвешенными в воде мельчайшими органическими частичками. Наиболее известны комары из родов Culex и Aedes (немалярийные комары) — переносчики возбудителей японского энцефалита, сибирской язвы, желтой лихорадки, а также Anopheles (малярийные комары) — специфические переносчики малярийного плазмодия. Доказано, что восприимчивость комаров к заражению возбудителями малярии определяется генотипически и наследуется моногенно. Малярийные и немалярийные комары легко отличаются друг от друга на всех стадиях их жизненного цикла (рис. 21.13). Яйца малярийных комаров р. Anopheles располагаются на поверхности воды поодиночке, и каждое снабжено двумя воздушными поплавками. Личинки их плавают в горизонтальном положении под поверхностью воды, а на предпоследнем членике имеют пару дыхательных отверстий. Куколки по форме напоминают запятые, находятся, как и личинки, под водной поверхностью и дышат кислородом воздуха через дыхательные рожки, имеющие форму широких воронок. Взрослые малярийные комары, сидя на предметах, располагаются под углом к их поверхности головкой книзу. Находящиеся по обе стороны от хоботка нижнечелюстные щупики равны ему по длине либо немного короче.
Рис. 21.13. Комары на разных стадиях жизненного цикла. А — Anopheles sp.; Б— Culex sp.: I — яйца, II — личинки. III — куколки, IV — имаго; 1— воздушные попалвки, 2— плотикиз яиц, 3— дыхательные отверстия, 4— дыхательный сифон, 5—воронкообразные дыхательныерожки, 6— цилиндрические дыхательные рожки, 7 — нижнечелюстные щупики
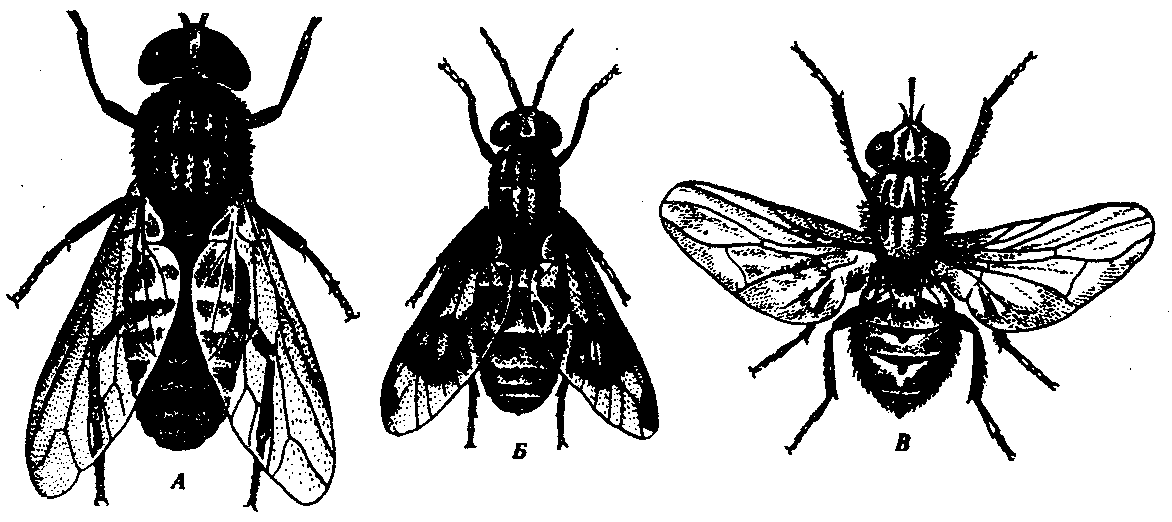
Немалярийные комары pp.Culex и Aedes откладывают яйца, слипающиеся группами в небольшие плотики серо-стального цвета. Личинки располагаются под поверхностью воды под углом к ней и имеют на предпоследнем членике длинный дыхательный сифон. Дыхательные рожки куколок имеют форму тонких цилиндрических трубочек, а нижнечелюстные щупики взрослых комаров коротки и достигают не более трети длины хоботка. Тело немалярийные комары держат параллельно поверхности, на которой сидят. Борьба с комарами наиболее результативна в отношении водных стадий жизненного цикла — личинок и куколок. Применяются мелиоративные методы — засыпка канав и карьеров со стоячей водой. Возможна обработка ядохимикатами отдельных водоемов с большой концентрацией личинок и куколок, а также мест массовых скоплений половозрелых стадий комаров в дневное время суток (сараи, скотные дворы). Наиболее эффективными являются биологические меры борьбы в сочетании с гидромелиоративными, проводящимися в соответствии с государственными антималярийными программами. Так, в Западном Закавказье удалось быстро снизить численность комаров и заболеваемость населения малярией за счет мелиорации и разведения рыб — гамбузий, питающихся преимущественно личинками двукрылых. Для индивидуальной защиты применяют репелленты и механические средства: марлевые пологи, сетки и т. д. 3.53: Мошки (сем. Simuliidae). Мелкие кровососущие насекомые длиной 2—6 мм. Ротовой аппарат короткий и очень мощный, предназначен для прокалывания кожи и слизывания крови. Крылья прозрачны, без пятен, конечности короткие и толстые (рис. 21.14, Б). Распространены повсеместно, но особую опасность представляют в Африке и тропической Америке, где являются переносчиками возбудителей онхоцеркоза. Интересно, что мошки кусают человека и животных преимущественно в бедра и боковые поверхности туловища. Именно в этих областях скапливаются микрофилярии онхоцерков, что значительно увеличивает вероятность их передачи. В большом количестве мошки встречаются поблизости от ручьев и речек с быстрым течением, чистой и прохладной водой. На дне таких водоемов самки откладывают яйца, из которых выходят личинки, ведущие прикрепленный и малоподвижный образ жизни. Через 2—3 недели формируется куколка, и еще через 1 неделю из ее оболочки выходят взрослые мошки. Развитие их идет синхронно, и огромное количество мошек на больших территориях появляются одновременно. Одним из наиболее экологически чистых методов борьбы с мошками является механическая очистка зон особенно быстрого течения рек и ручьев, где на дне и лежащих предметах скапливаются личинки мошек. При этом они отрываются от места прикрепления и уносятся течением. Многие из них при этом гибнут или поедаются различными хищниками. В редких случаях в очагах онхоцеркоза при высокой концентрации личинок мошек на небольших площадях возможно применение инсектицидов. Мокрецы (сем. Ceratopogonidae). Самые мелкие из кровососущих двукрылых. Их размеры от 1 до 4 мм. Большинство видов имеют пятнистые крылья, длинные многочленистые усики и колюще-сосущий хоботок (рис. 21.14, В). Обитают повсеместно, но так же, как мошки, в тропиках являются переносчиками возбудителей некоторых филяриатозов. Размножаются мокрецы в небольших стоячих водоемах или во влажной почве. Развитие от яйца до имагинальной стадии продолжается около 1 мес. На человека нападают преимущественно при высокой влажности воздуха или во время дождя. Против мокрецов наиболее эффективны индивидуальные средства защиты. Слепни (сем. Tabanidae) — наиболее крупные кровососущие двукрылые, до 30 мм длиной. Общим видом напоминают крупных мух. Ротовой аппарат сочетает в себе черты колюще-сосущего и лижущего.
Рис. 21.15. Крупные кровососущие двукрылые. А — слепень Tabanus sp.; Б — слепень Chrysophthalmus sp.; В — муха осенняя жигалка
Все тело слепней покрыто тонкими щетинками (рис. 21.15, А, Б). Распространены повсеместно. В северных широтах являются переносчиками бактерий — возбудителей сибирской язвы и туляремии, а в тропиках и филярий — возбудителей лоаоза. Американские слепни кусают человека и животных преимущественно в голову, где располагаются микрофилярии. Слепни — теплолюбивые и светолюбивые насекомые. Они активны в жаркие дневные часы на севере только летом, а в тропиках — в любом сезоне. На человека нападают обычно около воды. Яйца самки слепней откладывают на околоводных растениях. Личинки развиваются во влажном иле около уреза воды и ведут хищнический образ жизни. Цикл развития в среднем продолжается около 1 года. Против укусов слепней эффективны средства индивидуальной защиты. 3.54,3.55: Осенняя жигалка Stomoxys calcitrans — муха средних размеров серого цвета со сверлящим хоботком (рис. 21.15, В). Кровью питаются как самцы, так и самки. Личинки развиваются в навозе. В умеренных широтах массовая активность мух наблюдается в конце лета и начале осени, а в тропиках — круглый год. Держится обычно около животноводческих комплексов, активно нападает и на животных, и на человека. Является механическим переносчиком возбудителей туляремии и сибирской язвы. Для борьбы с массовым выплодом жигалок необходимо гигиеническое содержание домашних животных, а предохранение от укусов достигается индивидуальными средствами защиты. Мухи це-це р. Glossina- широко распространены в экваториальной Африке. Довольно крупные мухи длиной до 13,5 мм. Ротовой аппарат сходен с хоботком осенней жигалки (рис. 21.15, В). Самки живородящи, рождают периодически по одной личинке, которая сразу окукливается, углубляясь в почву. Через 3 недели появляется имагинальная форма. Размножение происходит в тени деревьев и кустов на берегах водоемов. Несколько близких видов мух це-це отличаются друг от друга особенностями окраски, а главное — биологии: одни виды поселяются преимущественно около жилищ человека и питаются в основном его кровью и кровью домашних животных. Другие — обитают в естественной природе (в саваннах и лесах), предпочитая питаться кровью крупных диких копытных, а человека кусают случайно.Все виды являются специфическими переносчиками возбудителя африканского трипаносомоза (см. разд. 19.3.2). Основной мерой борьбы является индивидуальная защита от укусов. Вольфартова муха Wohlfahrtia magnified - обитает в южных районах Европы, на Ближнем Востоке, в Средней Азии. Взрослые формы питаются нектаром цветов. Они рождают одновременно 150—190 живых личинок в кожные ранки и на неповрежденные слизистые оболочки крупных млекопитающих и человека. Личинки активно проникают в ткани хозяина, питаясь ими и производя серьезные повреждения. Через несколько суток личинки покидают хозяина и окукливаются в почве. Основная профилактика тканевых миазов — применение репеллентов при контакте со скотом, около которого обычно концентрируется большое количество двукрылых. 4.1: представление разнообразие возникновение живых форм в результате саморазвития природы уходят корнями в глубь веков. В древнеиндийских текстах Аюрвед, датируемых несколькими тысячелетиями до н.э, указывается что человек произошел от обезьяны. За два тысячелетия до н.э в Китае существовали учения, допускающие превращение одних организмов в другие путем их постепенных изменений. Мысль о развитии всего многообразия неодушевленных и одушевленных природных объектов путем превращения органического числа элементарных начал неоднократно высказывалась мыслителями древней Греции. Гераклит (конец 4 начало 5 века до н.э) рассматривал движение как основу мира. Эмпедокл (5 век до н.э) считал, что все существующее происходит из четырех начал, таких как огонь, земля, вода, и воздух, управляемых объединяющей и разъединяющей силами- любви и ненавистью., причем первыми из живых существ появились растения, а затем животные. Демокрит (5 век до н.э) отмечал приспособление отдельных органов к выполняемым функциям. Аристотель (384-322 годы до н.э) указывал на черты сходства и различия у отдельных живых существ и составил классификацию растений и животных. Ф.Энгельс назвал мысли древних философов «гениальными догадками». Одновременно он подчеркивал их умозрительный характер, поскольку зачастую они были плодом размышлений и не опирались на данные систематических наблюдений природы. Таким образом, представления мыслителей древности отличались незавершенностью и включали элементы очевидной фантазии. По этой причине было бы неправильно рассматривать античных мыслителей как предшественников современных эволюционистов. 4.2: К.Линней: создатель единой системы растительного и животного мира. Заложил основы современной биноминальной номенклатуры, введя в практику систематики двойные названия (родовое и видовое, например, Homo sapiens – человек разумный), которые позже стали использоваться в качестве видовых эпитетов в биноминальных названиях живых организмов. Линней К. разделил природный мир на три царства: минеральное, растительное и животное, используя четыре уровня (ранга): виды, роды, отряды и классы. Введенный Линнеем метод формирования научного названия для каждого из видов используется до сих пор. К. Линней первый обратил внимание на универсальность и реальность вида как структурной единицы живой природы, разработал: • правила классификации и номенклатуры организмов, • занимался поисками естественной системы растений и животных, • предложил первое научное определение вида, • уточнил критерии вида. Ш.Бонне: учение о наличии в половых клетках организмов материальных структур, предопределяющих развитие зародыша и признаки образующегося из него организма (все части организма заложены в половых клетках). На основе преформизма Ш. Бонне создал теорию вложения: в организме первичной самки каждого вида заложены все будущие поколения, а, следовательно, всякое развитие заранее предопределено. Теория вложения получила конкретное воплощение в «лестнице существ»: предустановленное и изначально заложенное развертывание Природы от низших форм к высшим формам. Теория вложения представляла собой распространение преформизма на развитие всего животного и растительного мира. Преформированное индивидуальное развитие обозначалось термином «эволюция» и после работ Ш. Бонне этот термин стал применяться к историческому развитию организмов. Ж.Бюффон: Жорж Бюффон одним из первых изложил концепцию ТРАНСФОРМИЗМА (лат. transformare превращать, преобразовывать) – представления об изменении и превращении форм животных и растений, ограниченная изменчивость видов, происхождение видов в пределах относительно узких подразделений под влиянием среды. Ж. Бюффон сформулировал идею единства живой природы и единства плана строения живых существ. Основные идеи Ж. Бюффона: · возникновение живых организмов из неорганических веществ; · развитие от простых форм к сложным; · единство происхождения животных организмов; · виды животных и растений изменяются под влияние внешней среды;· наследование приобретенных признаков. Ж. Бюффон видел доказательство единства происхождения в плане строения животных и объяснял сходство близких видов их происхождением от общих предков. Ж.Кювье: Исходя из особенностей строения нервной системы животных, Ж. Кювье сформулировал в 1812 г. учение о четырех «ветвях» (типах) организации животных: позвоночные, членистые, мягкотелые и лучистые, между которыми, однако, не признавал никаких связей и переходов В пределах типа позвоночных Кювье различал 4 класса: млекопитающие, птицы, земноводные (вместе с пресмыкающимися) и рыбы. Описал большое число ископаемых форм животных и выявил принадлежность многих из них (ихтиозавры, плезиозавры, мегалозавры, летающие ящеры и др.) к определённым слоям земной коры; предложил по ископаемым остаткам организмов определять возраст геологических слоев и наоборот. И хотя Ж. Кювье отвергал эволюционные представления своего времени, собранный им фактический материал послужил обоснованием эволюции живой природы. Ж.Сент-Илер: Основную часть своей жизни он посвятил разработке идеи единства типа животных. В работе «Философия анатомии» он изложил «теорию аналогов» (гомологов по позднейшей терминологии), положенную им в основу сравнения разных организмов. Второй том этого труда был посвящен тератологии – науке об уродствах, в частности, экспериментальному изучению уродств; ученому удалось показать, что различные «монстры» – аномалии индивидуального развития – тоже обнаруживают единство плана строения с нормальными организмами. В эволюционном процессе Сент-Илера придавал ведущее значение изменяющему действию внешней среды (условиям окружающей среды) на организмы и возникновению внезапных отклонений в зародышевом развитии. Современные виды животных появились постепенно, они развивались исторически из ранее существующих форм. Сент-Илер разделял идеи трансформизма и полагал, что одни виды могут превращаться в другие под действием меняющихся условий среды. Ж.Ламарк: стал первым биологом, который попытался создать стройную и целостную теорию эволюции живого мира. По широте научных взглядов Ламарк далеко превосходил своих предшественников – трансформистов. Наибольшую известность Ж.Б. Ламарку принесли биологические труды, им же был предложен термин «биология» в его современном смысле. Как ботаник Ж.Б. Ламарк приобрел известность, издав трехтомник «Флора Франции», в которой впервые был представлен определитель, построенный по дихотомическому принципу (общепринятому ныне). Ч.Дарвин: Таким образом, дарвиновская теория эволюции опирается на следующие принципы: 1) наследственности и изменчивости; 2) борьбы за существование; 3) естественного отбора. Эти принципы являются краеугольным основанием научной биологии. Э. Геккель называл Ч. Дарвина «Ньютоном органического мира». Символично, что в Вестминстерском аббатстве Ч. Дарвин похоронен рядом с И. Ньютоном. В этом видится сближение имен двух великих ученых.
Сущность дарвиновской концепции эволюции: 1) эволюируют не отдельные особи, а виды и популяции; 2) виды в природе ведут борьбу за существование между собой и с условиями среды обитания; 3) борьба за существование и естественный отбор на основе наследственной изменчивости – основные движущие факторы эволюции. Результатом борьбы за существование и естественного отбора являются: · приспособленность организмов к условиям среды; · дивергенция (развитие нескольких дочерних видов от общего предкового вида); · прогрессивная эволюция (усложнение и совершенствование видов). 4.3: материалистические тенденции в объяснении мира и идея развития нашли отражение в научной мысли России 18 века. Основоположником русской науки М.В.Ломоносов (1711-1765) утверждал первичность материи, определения ее сущности природных тел и явлений. В качестве основного свойства материи он назвал движение, понимаемое как развитие. Материалистичны идеи А.Н.Радищева (1749-1802), изложенные в тракте «о человеке, его смертности и бессмертии». Он писал, что бытие вещей независимо от сил познания о них существует само по себе и что в это бытие нельзя проникать иначе, как через опыт. В построенной им «лестнице веществ» А.Н.Радищев обходится без божественных чинов. Ступени «лестницы» соответствует значительным этапам развития природы- превращению неорганических веществ в органические, возникновению у живых существ новых качеств, таких как ощущение, мышление. Отражением идеи развития в эмбриологии была концепция эпигенеза К.Ф.Вольфа (1733-1794), который в противовес метафизической теории преформизма утверждал, что развитие зародыша идет от простого к сложному. Русский ученый К.Ф.Вольф обращал внимание на изменчивость животных и растений, причину которой он видел в действии климатических факторов и особенностях питания. 4.4: Естественный отбор: Различия между организмами по приспособленности, оцениваемой передачей аллелей следующему поколению, выявляются в природе с помощью - естественного отбора. Главный результат отбора заключается не просто в выживании более жизнеспособных, а в относительном вкладе таких особей в генофонд дочерней популяции. Необходимой предпосылкой отбора является борьба за существование — конкуренция за пищу, жизненное пространство, партнера для спаривания. Естественный отбор происходит на всех стадиях онтогенеза организмов. На дорепродуктивных стадиях индивидуального развития, например в эмбриогенезе, преобладающим механизмом отбора служит дифференциальная (избирательная) смертность. В конечном итоге отбор обеспечивает дифференциальное (избирательное) воспроизведение (размножение) генотипов. Благодаря естественному отбору аллели (признаки), повышающие выживаемость и репродуктивную способность, накапливаются в ряду поколений, изменяя генетический состав популяций в биологически целесообразном направлении. В природных условиях естественный отбор осуществляется исключительно по фенотипу. Отбор генотипов происходит вторично через отбор фенотипов, которые отражают генетическую конституцию организмов. Как элементарный эволюционный фактор естественный отбор действует в популяциях. Популяция является полем действия, отдельные особи — объектами действия, а конкретные признаки — точками приложения отбора. В зависимости от формы отбор сокращает масштабы изменчивости, создает новую или сохраняет прежнюю картину разнообразия. Как и другие элементарные эволюционные факторы, естественный отбор вызывает изменения соотношений аллелей в генофондах популяций. Особенность его действия состоит в том, что эти изменения направленны. Отбор приводит генофонды в соответствие с критерием приспособленности. Он осуществляет обратную связь между изменениями генофонда и условиями обитания, накладывает на эти изменения печать биологической целесообразности (полезности). Естественный отбор действует совместно с другими эволюционными факторами. Поддерживая генотипическое разнообразие особей в ряду поколений, мутационный процесс, а также популяционные волны, комбинативная изменчивость создают для него необходимый материал. Популяционные волны: Популяционными волнами или волнами жизни (С.С. Четвериков) называют периодические или апериодические колебания численности организмов в природных популяциях. Это явление распространяется на все виды животных и растений, а также на микроорганизмы. Причины колебаний часто имеют экологическую природу. Отмечаемое в этом случае увеличение кормовых ресурсов способствует росту численности хищников, что, в свою очередь, интенсифицирует истребление жертвы. В настоящее время существенно возросли размеры популяций домовой мухи, находящей прекрасную кормовую базу в виде разлагающихся пищевых отбросов вблизи поселений человека. При росте численности организмов наблюдается слияние ранее разобщенных популяций и объединение их генофондов. Так как популяции по своему генетическому составу уникальны, в результате такого слияния возникают новые генофонды с измененными по сравнению с исходными частотами аллелей. На гребне популяционной волны некоторые группы особей выселяются за пределы ареала вида и оказываются в необычных условиях существования. В таком случае они испытывают действие новых факторов естественного отбора. Повышение концентрации особей в связи с ростом их численности усиливает внутривидовую борьбу за существование. Популяционные волны — это эффективный фактор преодоления генетической инертности природных популяций. Вместе с тем их действие на генофонды не является направленным. В силу этого они, так же как и мутационный процесс, подготавливают эволюционный материал к действию других элементарных эволюционных факторов. Изоляция: Ограничение свободы скрещиваний (панмиксии) организмов называют изоляцией. Снижая уровень панмиксии, изоляция приводит к увеличению доли близкородственных скрещиваний. Сопутствующая этому гомозиготизация усиливает особенности генофондов популяций, которые создаются вследствие мутаций, комбинативной изменчивости, популяционных волн. Препятствуя снижению межпопуляционных генотипических различий, изоляция является необходимым условием сохранения, закрепления и распространения в популяциях генотипов повышенной жизнеспособности. В зависимости от природы факторов ограничения панмиксии различают географическую, биологическую и генетическую изоляцию. Географическая изоляция заключается в пространственном разобщении популяций благодаря особенностям ландшафта в пределах ареала вида — наличию водных преград для «сухопутных» организмов, участков суши для видов-гидробионтов, чередованию возвышенных участков и равнин. Ей способствует малоподвижный или неподвижный (у растений) образ жизни. Биологическая изоляция возникает вследствие внутривидовых различий организмов и имеет несколько форм. К экологической изоляции приводят особенности окраски покровов или состава пищи, размножение в разные сезоны, у паразитов — использование в качестве хозяина организмов разных видов. Длительная экологическая изоляция способствует дивергенции популяций вплоть до образования новых видов. Этологическая (поведенческая) изоляция существует благодаря особенностям ритуала ухаживания, окраски, запахов, «пения» самок и самцов из разных популяций. При физической (механической) изоляции препятствием к скрещиванию являются различия в структуре органов размножения или просто разница в размерах тела. У растений такая форма изоляции возникает при приспособлении цветка к определенному виду опылителей. Описанные формы изоляции, особенно в начальный период их действия, снижают, но не исключают полностью межпопуляционные скрещивания. Генетическая (репродуктивная) изоляция создает более жесткие, иногда непреодолимые барьеры скрещиваниям. Она заключается в несовместимости гамет, гибели зигот непосредственно после оплодотворения, стерильности или малой жизнеспособности гибридов. Мутационный процесс: Изменения наследственного материала половых клеток в виде генных, хромосомных и геномных мутаций происходят постоянно. Особое место принадлежит генным мутациям. Они приводят к возникновению серий аллелей и, таким образом, к разнообразию содержания биологической информации. Изменяя частоту одного аллеля по отношению к другому, он оказывает на генофонд популяции прямое действие. Еще большее значение имеет формирование за счет мутантных аллелей резерва наследственной изменчивости. Это создает условия для варьирования аллельного состава генотипов организмов в последовательных поколениях путем комбинативной изменчивости. Благодаря мутационному процессу поддерживается высокий уровень наследственного разнообразия природных популяций. Совокупность аллелей, возникающих в результате мутаций, составляет исходный элементарный эволюционный материал. В процессе видообразования он используется как основа действия других элементарных эволюционных факторов. Большинство мутаций первоначально оказывает на фенотип особей неблагоприятное действие. В силу рецессивности мутантные аллели обычно присутствуют в генофондах "популяций в гетерозиготных по соответствующему локусу генотипах. Благодаря этому достигается тройственный положительный результат: 1) исключается непосредственное отрицательное влияние мутантного аллеля на фенотипическое выражение признака, контролируемого данным геном; 2) сохраняются нейтральные мутации, не имеющие приспособительной ценности в настоящих условиях существования, но которые смогут приобрести такую ценность в будущем; 3) накапливаются некоторые неблагоприятные мутации, которые в гетерозиготном состоянии нередко повышают относительную жизнеспособность организмов (эффект гетерозиса). Таким образом создается резерв наследственной изменчивости популяции. Мутационный процесс, выполняя роль элементарного эволюционного фактора, происходит постоянно на протяжении всего периода существования жизни, а отдельные мутации возникают многократно у разных организмов. Генофонды популяций испытывают непрерывное давление мутационного процесса. Это обеспечивает накопление мутаций, несмотря на высокую вероятность потери в ряду поколений единичной мутации. Дрейф гены: Мутации и комбинативная изменчивость, периодические колебания численности организмов, изоляция изменяют генофонды популяций случайным образом. Их совместное действие с естественным отбором в процессе видообразования придает биологической изменчивости в целом приспособительный характер. Выполнению отбором упорядочивающей роли препятствуют изменения частот аллелей, зависящие от случайных причин. Таковыми в данном случае являются причины, обусловливающие преимущественное размножение генотипов вне связи с их приспособительной ценностью. Так как динамика частот аллелей в генофондах последовательных поколений носит статистический характер (см. закон Харди — Вайнберга), размах случайных колебаний этого показателя возрастает по мере снижения численности особей в популяции. Расчеты показывают, что при воспроизведении 5000 потомков родительской популяции с частотой некоего аллеля р = 0,50 колебания концентрации этого аллеля в 99,994% вариантов дочерних популяций в силу случайных причин (в отсутствие отбора по этому аллелю) не выйдут за пределы 0,48—0,52. Если же родительская популяция мала и воспроизводит 50 потомков, то размах случайных колебаний концентрации наблюдаемого аллеля в том же проценте вариантов дочерних популяций составит 0,30—0,70. Случайные, но не обусловленные действием естественного отбора колебания частот аллелей называют генетико-автоматическими процессами или дрейфом генов. Дрейф генов обусловливает утрату (р = 0) или закрепление аллелей в гомозиготном состоянии у всех членов популяции (р = 1) вне связи с их приспособительной ценностью. Он играет важную роль в формировании генофондов малочисленных групп организмов, изолированных от остальной части вида. 4.5: Наряду с общей приспособленностью, зависящей от генотипа в целом и измеряемой выживаемостью и успехом в размножении, в процессе эволюции возникают приспособления, или адаптации, для решения организмом экологических задач, предъявляемых средой обитания. Отдельные адаптации — это постоянно возникающие в процессе развития жизни, изменяющиеся, самосовершенствующиеся, иногда исчезающие, эволюционно обусловленные приспособления к конкретным факторам среды. В результате выработки адаптации достигается состояние адаптированности, или соответствия морфологии, физиологии, поведения организмов занимаемым ими экологическим нишам. Под экологической нишей понимают всю совокупность условий среды и образа жизни данного организма. Процесс выработки адаптации происходит постоянно. В него вовлечены многие признаки организма. Эволюция птиц от рептилий включала, например, последовательные изменения костей, мышц, покровов, конечностей. Увеличение грудины, перестройка гистологической структуры костей, придавшей им наряду с прочностью легкость, развитие оперения, обусловившего лучшие аэродинамические свойства и терморегуляцию, превращение пары конечностей в крылья, обеспечило решение проблемы полета. У некоторых представителей птиц впоследствии развились приспособления к наземному или водному образу жизни (страус, пингвин). Вторичные адаптации захватили также ряд признаков: пингвины, например, сменили крылья на плавники, а их покровы стали водонепроницаемы. Одна экологическая задача может решаться путем выработки разных адаптации. Так, средством термоизоляции у медведей, песцов является густой мех, а у китообразных — жировой подкожный слой. Адаптации возникают в ответ на конкретную экологическую задачу. В силу этого они всегда относительны. Относительность адаптации заключается в ограниченности их приспособительного значения определенными условиями обитания. Так, приспособительная ценность пигментированности бабочек березовых пядениц по сравнению со светлыми формами очевидна лишь на закопченных стволах деревьев. Приведенный пример показывает также, что о степени адаптивности признака можно судить лишь сравнив два его разных состояния. К адаптации иногда приводит новая мутация. Включившись в систему генотипа, она изменяет фенотип в направлении более эффективного решения экологических задач. Этот путь возникновения адаптации называют комбинативным. Есть несколько классификаций адаптации. По механизму действия выделяют приспособления пассивной защиты (высокая плодовитость; покровительственная, отпугивающая окраска), активной защиты, перемещения и добывания пищи (совершенствование аппарата движения, нервной системы, органов чувств; развитие средств нападения у хищных), к общественному образу жизни (разделение «труда» у пчел), к сожительству с другими организмами (симбиоз, паразитизм). В зависимости от характера изменения различают адаптации с усложнением или упрощением морфофизиологической организации. Последнее типично для паразитов. По масштабу приспособления делят на специализированные и общие. С помощью специализированных адаптации организм решает конкретные задачи в узкоограниченных условиях жизни вида. К примеру, особенности строения языка муравьеда таковы, что обеспечивают питание муравьями. Общие адаптации позволяют решать многие задачи в широком спектре условий среды. К ним относят внутренний скелет позвоночных и наружный членистоногих, гемоглобин как переносчик кислорода и др. 4.6: Размножение человека осуществляется половым путем, а репродуктивные ареалы в той или иной степени ограничены определенной группой населения. Это позволяет выделить в человечестве сообщества, аналогичные популяциям в биологическом понимании этого термина. В антропогенетике популяцией называют группу людей, занимающих общую территорию и свободно вступающих в брак. Изоляционные барьеры, препятствующие заключению брачных союзов, нередко носят выраженный социальный характер (например, различия в вероисповедании). Благодаря этому в формировании популяций людей главную роль играет не общность территории, а социальные факторы. Размер, уровень рождаемости и смертности, возрастной состав, экономическое состояние, уклад жизни являются демографическими показателями популяций людей. Генетически они характеризуются генофондами (аллелофондами). Демографические показатели оказывают серьезное воздействие на состояние генофондов человеческих популяций, главным образом через структуру браков. Большое значение в определении структуры браков имеет размер группы. Популяции из 1500—4000 человек называют демами, популяции численностью до 1500 человек — изолятами. Для демов и изолятов типичен относительно низкий естественный прирост населения — соответственно порядка 20% и не более 25% за поколение. Частота внутригрупповых браков в них со
|
||
|
|
Последнее изменение этой страницы: 2016-08-16; просмотров: 1299; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.169 (0.029 с.) |