Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Нарушения структуры хромосомСодержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
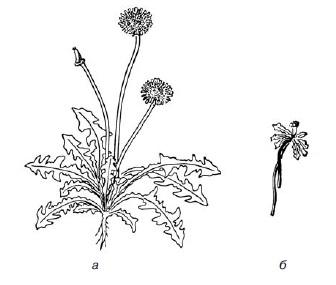
2.48. Изменчивость, её виды. Характеристика модификационной изменчивости, примеры Изменчивость организмов проявляется в разнообразии особей (одного вида, породы или сорта), отличающихся друг от друга по комплексу признаков, свойств и качеств. Причины тому могут быть разными. В одних случаях данные различия (при одинаковых генотипах у организмов) определяются условиями среды, в которых происходит развитие особей. В других — различия обусловлены неодинаковыми генотипами организмов. На основании этого выделяют два типа изменчивости: ненаследственную (модификационную, фенотипическую) и наследственную (генотипическую). Модификационная (фенотипическая) изменчивость заключается в том, что под действием разных условий внешней среды у организмов одного вида, генотипически одинаковых, наблюдается изменение признаков (фенотипа). Изменения эти индивидуальны и не наследуются, т. е. не передаются особям следующих поколений. Рассмотрим проявление подобной закономерности на нескольких примерах. В одном из опытов корневище одуванчика разрезали вдоль острой бритвой и высадили половинки в разных условиях — в низине и в горах. К концу сезона из этих проростков выросли совершенно не похожие друг на друга растения. Первое из них (в низине) было высоким, с большими листьями и крупным цветком. Второе, выросшее в горах, в суровых условиях, оказалось низкорослым, с мелкими листьями и цветком (рис. 1). Генотип у этих двух растений абсолютно идентичен (ведь они выросли из половинок одного корневища), но их фенотипы существенно различались в результате разных условий произрастания. Потомки этих двух растений, выращенные в одинаковых условиях, ничем не отличались друг от друга. Следовательно, фенотипические изменения не наследуются.
Рис. 1. Изменение одуванчика под влиянием внешних условий среды (по Боннье): à — растение, выращенное в низине; á — в горах; оба растения — отводки одной особи Биологическое значение модификационной изменчивости заключается в обеспечении индивидуальной приспособляемости организма к различным условиям внешней среды. Рассмотрим другой пример. Представим себе, что два брата, однояйцовых близнеца (т. е. с идентичными генотипами) выбрали еще в детстве разные увлечения: один посвятил себя тяжелой атлетике, а другой — игре на скрипке. Очевидно, через десяток лет между ними будет наблюдаться существенное физическое различие. И также ясно, что у спортсмена его новорожденный сын не родится с «атлетическими» признаками. Изменения фенотипа под воздействием условий внешней среды могут происходить не беспредельно, а только в ограниченном диапазоне (широком или узком), который обусловлен генотипом. Диапазон, в пределах которого признак может изменяться, носит название нормы реакции. Так, например, признаки у коров, учитываемые в животноводстве, — удойность (т. е. количество вырабатываемого молока) и жирность молока — могут изменяться, но в разных пределах. В зависимости от условий содержания и кормления животных удойность варьируется существенно (от стаканов до нескольких ведер в сутки). В данном случае говорят о широкой норме реакции. А вот жирность молока очень незначительно колеблется в зависимости от условий содержания (всего на сотые доли процента), т. е. этот признак характеризуется узкой нормой реакции. Итак, условия внешней среды обусловливают изменения признака в пределах нормы реакции. Границы же последней продиктованы генотипом. Следовательно, изменения самой нормы реакции могут произойти только в результате изменения генотипа (т. е. в результате генотипической изменчивости). 2.49. Комбинативная изменчивость и её механизм Комбинативная изменчивость имеет две основные составляющие; 1) случайное, равновероятное расхождение хромосом в мейозе (оно обеспечивает перекомбинацию родительских хромосом и служит цитологическим обоснованием закона свободного комбинирования, сформулированного Г. Менделем) и 2) рекомбинация сцепленных генов, локализованных в гомологичных хромосомах. В более узком смысле под рекомбинацией подразумевают перекомбинацию генов, и потому предпосылкой для нее, в частности, и для комбинативной изменчивости вообще, является гетерозиготность организма по одному или более генам. Эта гетерозиготность, а следовательно, и рекомбинация возникают у эу- и прокариот разными путями: для их реализации у прокариот существуют конъюгация, трансформация и транедукция, а также — совместная инфекция (у вирусов). У эукариот гетерозиготность обеспечивается диплоидностью генома, а сама рекомбинация может происходить как в половых, так и в соматических клетках. Результатом рекомбинации в конечном итоге является перенос участков ДНК с одной молекулы на другую. В случае реципрокной рекомбинации этот перенос - взаимный, а при нереципрокной - односторонний. Существуют два подхода к изучению процесса рекомбинации. Первый из них, классический, анализирует наследование признаков и, если признаки имеют тенденцию наследоваться совместно, оценивает степень их сцепления, или частоту рекомбинации между соответствующими локусами. Этот подход возник в «домо-лекулярное» время и представляет собой статистический анализ наблюдаемого расхождения признаков при передаче их последующим поколениям. Второй подход к изучению генетической рекомбинации, молекулярный, направлен на анализ тонких механизмов этого процесса. Хотя для обоих подходов предметом исследования является один и тот же процесс, само понятие генетической рекомбинации неоднозначно. Можно выделить три типа рекомбинации:
Генетическая рекомбинация. Примером является мейотическая рекомбинация (кроссинговер) у эукариот, которая происходит в клетках после репликации, в профазе первого мейотического деления. Во время лептотены хромосомы конденсируются и становятся видимыми, В каждой из них после репликации дуплексная ДН К представлена двумя сестринскими хроматидами. Под электронным микроскопом видно, что на стадии лептотены пара сестринских хроматид каждой хромосомы формирует единый осевой элемент. Установлено, что у млекопитающих он состоит из белков SCP2 и SCP3 (от англ. synaptonemal complex proteins). На следующей стадии, зиготене, гомологичные хромосомы начинают соприкасаться друг с другом (конъюгировать) на отдельных, пока еще коротких участках. Одновременно осевые элементы гомологичных хромосом начинают соединяться попарно с помощью белка SCP1, который протягивается поперек между ними в виде субмикроскопических волокон (филамент). По завершении конъюгации, на стадии пахитены гомологичные хромосомы оказываются объединенными в биваленты по всей длине за счет специфической структуры, состоящей из двух продольных белковых тяжей. Это - так называемые латеральные элементы, в состав которых входят осевые элементы с прикрепленными к ним, петлеобразно уложенными фибриллами хроматина сестринских хроматид. Латеральные элементы соединены между собой поперечными белковыми волокнами, которые в совокупности формируют третью продольную структуру - центральный элемент. Из двух латеральных и одного центрального элемента образуется электроноплотная трехполосная структура, так называемый, синаптонемный комплекс, в котором гомологичные хромосомы прилегают к латеральным элементам с двух сторон, и этот контакт происходит «точечно» (в местах прикрепления петель к синаптонемному комплексу по всей era длине). Функциональное значение этой структуры, напоминающей застежку «молнию», состоит в том, что, с одной стороны, она не дает конъюгирующим хромосомам необратимо слипнуться, а с другой стороны — закрепляет их в строго гомологичном относительно локализованных на них генах взаиморасположении. В зависимости от размера генома у разных видов могут варьировать размеры синаптонсмного комплекса: общая ширина его трехполосной ленты составляет от 76 до 240 нм, а длина соответствует также видоспецифичной длине бивалентов в профазе 1 мейоза. На стадии диплотены гомологичные хромосомы бивалентов начинают расходиться, но обнаруживается, что несестринские хроматиды в биваленте остаются сцепленными в некоторых точках, образуя фигуру, получившую название хиазмы. На стадии диакинеза хромосомы конденсируются путем спирализации, а хиазмы вследствие отталкивания гомологов начинают сдвигаться к краям хромосом. В этот момент все четыре хроматиды становятся видимыми. Это — прямые наблюдения, и они позволяют сделать некоторые предположения о процессе рекомбинации. Профаза первого деления мейоза — единственный момент, когда гомологичные хромосомы образуют комплекс друг с другом, что, является условием, необходимым для осуществления рекомбинации. Можно полагать, что именно в обеспечении рекомбинации и состоит суть синапса—образование синаптонемногокомгаткса, временной структуры, которая формируется на стадии зиготены и разрушается в диплотене. Согласно мнению авторов приведенной выше гипотетической схемы, синаптонемный комплекс «...необходим для организации хроматина ввиде серии латеральных петель, основания которых собраны в линейную последовательность на поверхности его латеральных элементов и доступны для узнавания гомологичных локусов и кроссинговера». Существует очень правдоподобная, но до настоящего времени не всеми исследователями разделяемая гипотеза, что хиазмы представляют собой места прохождения рекомбинаций- ведь количество тех и других примерно совпадает. Это позволяет локализовать время происхождения процесса рекомбинаций, и считать, что даже на стадии диакинеза в местах рекомбинации хромосомы все еще остаются связанными посредством нитей ДНК. 2.50. Генотипическая изменчивость. Хромосомные аберрации, примеры Наследственная (генотипическая) изменчивость составляет часть общей (фенотипической) изменчивости, которая определяется генетическими различиями между особями или группами особей. Ведущую роль в процессах генотипической И. играет изменение генотипа в результате изменения репродуцирующихся структурных элементов ядра (ДНК хромосом) — мутации. Генотипы являются не чем иным, как комбинациями генов. Поэтому биологическое значение полового размножения заключается именно в том, что образование разнообразных генотипов при таком размножении значительно ускоряется. Наряду с мутациями источником генотипической И. являются рекомбинации генов (так называемая рекомбинационная И.). На степень генотипической И. в популяции влияет также приток генов из других популяций. Изменения генотипа приводят к изменению характера развития того или иного признака в данных условиях окружающей среды, изменению наследственной нормы реакции и появлению различных отклонений (наследственные аномалии развития). Сходство врожденных пороков развития, возникающих в результате действия повреждающих факторов окружающей среды, и наследственных аномалий объясняется тем, что при мутациях и под влиянием тератогенов в клетках формирующихся тканей и органов нарушаются одни и те же звенья биохимических процессов. Хромосомные аберрации (хромосомные мутации, хромосомные перестройки) — тип мутаций, которые изменяют структуру хромосом. Классифицируют делеции (утрата участка хромосомы),инверсии (изменение порядка генов участка хромосомы на обратный), дупликации (повторение участка хромосомы), транслокации (перенос участка хромосомы на другую), а также дицентрические и кольцевые хромосомы. Известны такжеизохромосомы, несущие два одинаковых плеча. Если перестройка изменяет структуру одной хромосомы, то такую перестройку называют внутрихромосомной (инверсии, делеции, дупликации, кольцевые хромосомы), если же двух разных, то межхромосомной (дупликации, транслокации, дицентрические хромосомы). Хромосомные перестройки подразделяют также на сбалансированные и несбалансированные. Сбалансированные перестройки (инверсии, реципрокные транслокации) не приводят к потере или добавлению генетического материала при формировании, поэтому их носители, как правило, фенотипически нормальны. Несбалансированные перестройки (делеции и дупликации) меняют дозовое соотношение генов, и, как правило, их носительство сопряжено с клиническими отклонениями от нормы. Хромосомные перестройки играют определенную роль в эволюционном процессе и видообразовании, в нарушении фертильности, в онкологических и врождённых наследственных заболеваниях человека. Хромосомные перестройки были открыты у дрозофил при помощи генетического анализа. В некоторых скрещиваниях соотношение числа потомков в разных классах сильно отличалось от ожидаемого, и это объяснили наличием перестроек в хромосомах родителей. Делеции, дупликации и транслокации обнаружил К. Бриджес в 1916, 1919 и 1923 годах, соответственно. Первую инверсию описал А. Стёртевант в 1921 году, сравнивая порядок генов в хромосоме 3 у D.melanogaster и D.simulans. Первые наблюдения хромосомных перестроек были сделаны на политенных хромосомах слюнных желез. Лишь спустя некоторое время существование перестроек было доказано цитологически на митотических хромосомах. Однако проще всего перестройки можно увидеть в политенных хромосомах у гетерозиготных особей, благодаря образованию петель и крестообразных структур. Также перестройки можно увидеть в профазе мейоза при образовании синаптонемных комплексов, где, благодаря синапсису гомологичных хромосом, также образуются петли и крестообразные структуры.
|
||||
|
|
Последнее изменение этой страницы: 2016-08-16; просмотров: 909; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.14.247.170 (0.013 с.) |