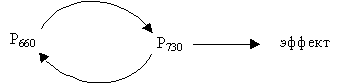Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Зависимость роста растений от экзогенных факторовСодержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
Рост, являясь интегральной функцией растения, зависит от влияния абиотических факторов внешней среды: света (его интенсивности, спектрального состава, продолжительности и периодичности), температуры (величины и периодичности), питательных веществ (количества и сбалансированности), механических воздействий ветер и др.). Регуляция роста светом. Разносторонние процессы, которые определяют влияние света на форму растений, были названы фотоморфогенезом. Растения, выросшие в темноте, называются этиолированными. Они отличаются от растений, выращиваемых на свету, рядом морфологических и биохимических особенностей. Программа развития нормального зеленого растения контролируется специальными световыми реакциями, лежащими в основе фотоморфогенеза. Хлорофилл и разные вспомогательные пигменты фотосинтетического аппарата не играют в этих процессах главной роли, посколько начальные этапы развития растений на свету не могут осуществляться за счет фотосинтеза: структурная и биохимическая основы фотосинтетического аппарата в этот период еще не сформированы. Для устранения эффекта этиоляции и включения процессов фотоморфогенеза достаточно ежедневного 5–10 минутного освещения растений. Термин «фотоморфогенез» объединяет процессы, которые не зависят от направления и периодического освещения, а определяются длиной волны и интенсивностью светового потока. Вероятно, в процессах фотоморфогенеза растений играют роль определенные соединения – пигменты. Содержание этих пигментов в растениях очень малое, поэтому количество энергии, необходимое для насыщения соответствующих фотопроцессов, на несколько порядков ниже, чем при фотосинтезе. В то время как при фотосинтезе 8–10 квантов света необходимо для выделения только одной молекулы О2, такое же количество квантов на одну клетку может полностью определить репродуктивную судьбу растения или направление роста всего стебля. Реакция растений на длительность светового периода (или длину дня) разная. Эта реакция растений на продолжительность дня получила название фотопериодизма. Выделяют растения короткого дня, когда растения зацветают при небольшой продолжительности освещения – менее некоторой критической величины. Когда же длина дня превышает определенную критическую величину – это растения длинного дня. Имеется и группа растений, цветение которых не зависит от длины дня – нейтральные растения. Критический фотопериод у разных видов и даже сортов растений как короткого, так и длинного дня значительно варьирует. Эти различия в критическом фотопериодизме играют важную роль в размещении растений на Земле. После открытия фотопериодизма выявили, что многие растения реагируют на длительность непрерывной темноты, а не на протяженность светового периода. Иными словами, так называемые растения короткого дня – это на самом деле растения длинной ночи. Для закладки цветковых бугорков им необходимо определенная минимальная длительность темноты, которая не прерывается светом. Подобным образом растение длинного дня может быть в действительности растением короткой ночи: оно будет цвести только в том случае, если ночной период не продолжительней некоторого максимума. Всего только один подходящий период темноты может привести растение короткого дня к цветению, даже тогда, когда последующие темновые периоды недостаточно длительны. Этот феномен известен как фотопериодическая индукция. У многих растений длинного дня имеет место подобный, но обратно направленный феномен: прерывание очень длительного темнового периода вспышкой света приводит к индукции и заложению цветков. Последнее говорит о том, что растения длинного дня и короткого дня, вероятно, владеют одинаковыми фотопериодическим механизмом, но последний каким-то образом действует в разных направлениях. Это нашло дальнейшее подтверждение при изучении действия света разной длины волны – спектр действия. Для ингибирования цветения короткодневных растений и для активации цветения длиннодневных растений во всех случаях был определен максимум активности в красной области спектра (около 660 нм) при почти полной неэффективности других длин волн. Подобие спектров позволило считать, что зацветание растений как короткого, так и длинного дня контролируется одним и тем же пигментом. Далее, свет сильно влияет на прорастание семян. Некоторые семена плохо прорастают в полной темноте, но быстро и хорошо при экспонировании на свету на протяжении нескольких минут. Оказалось, что спектр действия для этого эффекта подобен спектру действия для цветения. Проросток (гороха), который выращивается в полной темноте, имеет очень длинный тонкий стебель, апикальный изгиб и почти не распустившиеся листья. Когда такой этиолированный проросток выставить только на короткое время на свет, листья после этого разворачиваются, изгиб начинает выпрямляться, удлинение стебля замедляется. Поэтому, появилась возможность заключить, что такие разносторонние реакции как прорастание семян, изменение этиолированного проростка (его деэтиолирование) и индукция цветения регулируются одним и тем же пигментом – рецептором. Как определили, таким пигментом является фитохром. Фитохром – от греческого слова «растение» и «краситель». Фитохром представлен растворимыми в воде соединениями, относящимися к билихромпротеинам. Как и у фикобилинпротеинов (фикоцианобелин), хроматофор (пигментная часть молекулы) фитохрома представлен незамкнутыми порфиринами (тетрапирол). Другая часть молекулы образуется бесцветным белком. После поглощения фитохромом света с определенной длинной волны форма хроматофора изменяется, и, это в свою очередь, изменяет форму белкового компонента. Молекулярная масса фитохрома колеблется в зависимости от вида растения от 120 до 170 кДа. В составе фитохрома – одна полипептидная цепь. Существует, как предполагают, фитохром в клетке в виде димера. Фитохром регулирует многие физиологические реакции, наблюдаемые при поглощении малых доз красного и дальнего красного света растениями, которые находились в полной темноте. Фитохром находится в растениях в двух формах: Р660 – поглощает красные лучи и превращается в Р730; последний поглощает дальние красные лучи и переходит в Р660 (рис. 6.16). Р660 (Рк) – физиологически неактивный.
Рис. 6.16. Схема биологического ответа при действии красного и дальнего красного света Структура фитохрома представлена на рис. 6.17.
Рис. 6.17.Структура фитохрома Реакции, управляемые фитохромом, зависят от концентрации Р730 (Рдк). Полагают, что Ф660 содержит на один протон больше, чем Ф730 (см. рис. 6.17). Эта разница влияет на конформацию хромотофора и белковой молекулы, с которой он связан. Фитохром присутствует в растениях, как уже отмечалось, в небольших количествах: его концентрация составляет около 1 мкМ. При этом он размещен в растении неравномерно, больше всего его в растущих, делящихся тканях, например в клетках меристемы. В клетках стебля он практически отсутствует. До освещения красным светом фитохром обычно распределен по всей цитоплазме и ее мембранах, а после короткого освещения выявляется только в определенных частях клетки. Вероятно, эти части – ни ядра, ни пластиды, ни митохондрии, а, скорее, ЭР, разбросанный по всей цитоплазме. Длительное воздействие света приводит к появлению фитохрома уже и в ядрах. Подтверждают эти выводы эксперименты с изолированными субклеточными фракциями; однако, не все согласны с интерпретацией полученных данных. Все же данные физиологических опытов позволяют предполагать, что фитохром может находиться в клетке во многих местах в соответствии с его многочисленными функциями. Вероятно, он локализован внутри этиопластов и митохондрий, или в их наружных мембранах, так как изолированные органеллы реагируют на воздействие красным и дальним красным светом, после их выделения. Предполагают, что фитохром находится (или каким-то образом связан) в плазмалемме. Считают даже, что хроматофору фитохрома свойственна определенная ориентация в плазмалемме. Кроме того, вероятно, что фитохром изменяет свою ориентацию в мембране на 90о при фототрансформации Обычно реакции достигают насыщения, когда 50 % фитохрома представлена формой Р730, и продолжается до той поры, пока имеется достаточное количество Р730. Превращение Р660 ↔ Р730 происходит также и in vitro. Как считают, оно связано с конформационными изменениями участков белковой части молекулы фитохрома. Этот процесс происходит через промежуточные этапы, потому что пигмент исчезает быстрее, чем появляется другой. Из-за того, что Р730 неустойчив, его количество в темноте уменьшается, что объясняется медленным тепловым превращением в Р660 и довольно быстрым, по всей вероятности, окислительным разрушением, катализируемым металлами. Очень маленькое количество Р730остается при выдерживании растений в темноте.
Рис. 6.18.Схема превращения фитохрома Потери фитохрома компенсируются invivo его синтезом, который начинается по принципу обратной связи, когда суммарное количество фитохромов (Р660 + Р730) становится ниже порогового (рис. 6.18). Синтез пигмента (незамкнутого порфирина) происходит, как и синтез хлорофилла (циклического порфирина), через δ-аминолевулиновую кислоту. Под влиянием Дк света Р730 быстро и почти целиком (около 90 %) превращается в Р660, а при красном свете Р660 превращается в Р730 примерно на 80 %. Солнечный свет содержит приблизительно одинаковые доли К и Дк. Это приводит к равновесию Р730/Р660, при котором содержание Р730 доходит до 50 % (во всяком случае, больше, чем в темноте). Поэтому в реакциях, регулируемых фитохромом, солнечный свет действует как красный. Важная роль фитохрома обусловлена тем, что это единственный из растительных пигментов, который использует световые сигналы для управления развитием растения. Ряд ферментативных реакций, связанных с процессом фотосинтеза регулируются фитохромом. В соответствии с общими представлениями, впервые высказанными Х. Мором (1966 г.), действие фитохрома связано в первую очередь с регуляторным влиянием Дк-формы на активность так называемых фотоиндуцибильных генов. В общем виде генетический путь трансдукции может быть представлен следующей схемой: К → Дк → активация или репрессии гена → рост или снижение содержания РНК → модификация содержания белкового продукта → биологический эффект. Кроме того, на основании полученных данных считают, что Са2+ и кальмодулин принимают участие в передаче сигнала от Дк-формы фитохрома к генетическому аппарату, а так же в этом процессе участвует и G-белок. Генетический механизм обеспечивает у растений медленные фитохромные ответы. Однако, быстрые фитохроминдуцируемые эффекты (фотоиндуцируемые изменения потенциала, набухание хлоропластов и т. д.), вероятно связаны с модификацией мембранных структур. В механизмах мембранной трансдукции фитохромного сигнала важную роль играет Са2+. Мембранный механизм передачи фитохромного действия имеет следующий вид: К → Дк → изменение проницаемости мембраны для ионов Са2+ → временное изменение концентрации Са2+ в цитоплазме → включение кальций-зависимой биохимической системы → биологический ответ. Кальциевый поток действует на активность кальмодулина, последний изменяет активность ферментов, например, протеинкиназ. Через действие катаболических ферментов может происходить активация фитогормонов, запускающих механизм ростовой фотореакции. Так состояние фитохрома может регулировать качественный и количественный состав гормонов четырех типов – этилена, цитокининов, ауксина и гиббереллинов – в разных органах растения. Кроме того, длительное облучение иногда контролирует уровень АБК. В некоторых случаях внесение гормона может снижать эффект трансформации фитохрома (например действие гиббереллина при деэтиляции). В природе фитохром, вероятно, контролирует форму растений и прорастание семян под пологом листвы, так как листья очень сильно поглощают свет в области 660 нм и относительно слабо – 730 нм. Фитохром может регулировать прорастание семян, деэтилирование, цветение, опадение листьев и даже старение. Синий свет также регулирует фотоморфогенетические реакции, в частности движение устьиц, фототропизм, ранние этапы синтеза хлорофилла и каротиноидов, движение листьев за солнцем, фототаксис хлоропластов, угнетение роста гипокотилей и стеблей в длину. Фоторецепторы, участвующие в восприятии синего света, называют криптохромами. У растений арабидопсиса выявлены соединения (криптохром 1, криптохром 2 и фототропин), которые воспринимают и передают сигналы синего света. Криптохропы – рецепторы синего света, локализованные в ядре и цитоплазме, включающие белковую и хромофорную составляющие. Функции хромофоров у криптохромов выполняют флавины (ФАД, деазафлавин) и (или) птерин. Криптохромная система замедляет рост гипокотеля (или междоузлия) на свету, помогает фитохрому контролировать процессы диэтиоляции. Обе фоторецепторные системы (криптохром и фитохром), действующие в ядерном компартменте, дополняют друг друга, что сказывается в интеграции информации о физиологически важных областях спектра – синей и красной. Фототропин представляет собой ассоциированный с мембраной белок, т. е. является мебранным рецептором синего света, функцию хромофора у которого выполняет флавин ФМН. Фототропин ответственен за фототропизм, хемотропизм и потенциалзависимые мембранные процессы. Характер ответных реакций растений при освещении синим светом определяется его интенсивностью и длиной волны. У папоротника Adiantumcapillus-veneris был найден ген, гомологичный как фитохромам, так и фототропинам. «Двойной» фоторецептор был назван суперхромом. Как и фототропин, суперхром обладает протеинкиназной активностью. Следовательно, если у одних растений фототропин взаимодействует с фитохромами опосредованно, то у Adiantum проблема взаимодействия красного и синего световых сигналов решена на уровне «двойного» рецептора. Таким образом, конечная форма зеленого растения, обусловленная в целом его генами, определяется участием света, который действует через разные фоторецепторы. Фитохром или сходный с ним пигмент имеется у зеленых водорослей, мхов, печеночников, папоротников, голосеменных и покрытосеменных, но, вероятно, отсутствует у грибов. Различные функции, которые он выполняет у этих разнородных организмов, свидетельствуют о его значительной адаптивной способности. Рассмотрим несколько примеров. Многим семенам из-за потребности в Фдк для прорастания необходим свет: под слоем почвы они остаются в покое, но хорошо прорастают на ее поверхности. Таковы, например, семена обычного сорняка Verbascum brattaria. В 1973 г., через 90 лет после пробного закапывания, 20% семян этого растения все еще были жизнеспособны и проростки по своему росту не отличались от контрольных, полученных из свежих семян. Хотя в течение долгого периода покоя такие семена продолжают дышать, что уменьшает резерв питательных веществ, расход последних может быть очень незначительным, если фитохром представлен формой Фк. Свежим семенам многих сорняков для прорастания свет не нужен, но потребность в свете возникает при помещении их в почву, особенно в переувлажненную. По-видимому, эти семена первоначально содержат много Фдк, который в темноте превращается снова в Фк. Этой реверсии не происходит, если семена хранятся в сухом воздухе, так как фитохром семян в этих условиях дегидратируется, а сухие Фк и Фдк не способны к взаимопревращению. Но если почва настолько влажная, что фитохром семян частично гидратируется, то происходит реверсия, которая у хранящихся семян, видимо, служит эффективным механизмом выживания. Некоторые виды семян прорастают глубоко в почве, куда свет не может проникать. Поскольку фитохром синтезируется в форме Фк, проростки на ранних этапах роста, когда они живут за счет запасных веществ эндосперма, содержат Фк, а не Фдк. У этиолированных, проростков бывают тонкие, быстро вытягивающиеся побеги с искривленным подсемядольным коленом и маленькие листья. Такой тип роста имеет значение для выживания, так как он позволяет проросткам быстро и экономно дотянуться.до света. Нежная верхушечная точка роста защищена изгибом, и энергия не растрачивается на рост листьев, поскольку они не могут функционировать до тех пор, пока растение не окажется на свету. Когда проросток выбьется через поверхность почвы на свет, некоторая часть фитохрома Фк превращается в ФдК. Изгиб затем выпрямляется, стебель становится толще, скорость его удлинения уменьшается, листья увеличивают свою поверхность и зеленеют, повышая фотосинтетическую активность. Таким образом, многие особенности проростка, существенные для выживания, регулируются превращением Фк. Рост под пологом листвы Свет, проходя через зеленый лист, в результате избирательного поглощения волн определенной длины хлорофиллом относительно обогащается лучами дальней красной области по сравнению с красной. Это препятствует прорастанию семян, чувствительных к свету, после развития листового полога. Поскольку прорастание в такой среде с недостатком света приводило бы к плохому росту, подавление прорастания, обусловленное отсутствием Фдк, имеет значение для выживания. Следующей весной перед развитием листового полога семена смогут прорасти и проростки будут хорошо развиваться. Затенение зелеными листьями влияет и на рост проростков. Поскольку фитохром в проростках нижнего яруса находится главным образом в форме Фк, рост затененных листовым пологом растений в некоторой мере сходен с ростом этиолированных проростков под землей: листья у них маленькие и тонкие, стебли необычно вытянуты. Поскольку они тратят значительную часть своих энергетических ресурсов на процессы, ведущие к удлинению стебля, растения нижнего яруса повышают свою способность достичь листового полога и проникнуть сквозь него; это известное преимущество в конкуренции, так как при затенении они получают очень мало лучистой энергии для фотосинтеза. По-видимому, не случайно пики поглощения Фк и хлорофилла а совпадают, в то время как пик поглощения Фда попадает в «окно» между короткими волнами, которые поглощаются фотосинтетическими пигментами, и длинными волнами, поглощаемыми водой, содержащейся в растительных тканях. Природа использовала эту ситуацию многочисленными сложными способами! 34. Клетка – наименьшая структура, обладающая всеми критериями живого: она растет, развивается, размножается и передает по наследству признаки, реагирует на внешние раздражители и способна к движению. Усилиями ученых М.Шлейдена и Т. Шванна в 1838-1839 гг. была создана клеточная теория, дополненная Р.Вирховым. В настоящее время клеточная теория включает в себя следующие положения: 1. Клетка – элементарная единица живого, способная к самообновлению, саморегуляции, самовоспроизведению и являющаяся единицей строения, функционирования и развития живых организмов. 2. Клетки всех живых организмов сходны по составу, строению и основным проявлениям жизнедеятельности. 3. Размножение клеток происходит путем деления исходной материнской клетки. 4. В многоклеточном организме клетки специализируются по функциям и образуют ткани, из которых построены органы и их системы, связанные между собой межклеточными, гуморальными и нервными способами регуляции. Различают два основных типа организации клеток: прокариотический и эукариотический. Прокариотические клетки наиболее просто организованы, не имеют обособленного ядра (табл. 1). К прокариотам относятся архебактерии, эубактерии, цианобактерии (сине-зеленые водоросли). Эукариотические клетки представляют более высокий тип клеточной организации, они имеют обособленное ядро и представлены клетками растений, грибов и животных. Сравнительная характеристика прокариотических и эукариотических клеток Таблица 1
* - коэффициент седиментации, указывает на скорость осаждения при ультрацентрифугировании, зависит от молекулярной массы и формы частиц.
Строение эукариотической клетки. Типичная эукариотическая клетка состоит из трех компонентов: цитоплазматической мембраны (плазмалеммы), цитоплазмы и ядра (рис. 12). Плазмалемма – двойной слой фосфолипидов с встроенными в него белками. Мембрана выполняет важные и весьма разнообразные функции: определяет и поддерживает форму клетки; защищает клетку от механических воздействий, проникновения повреждающих биологических агентов; осуществляет рецепцию многих молеку-лярных сигналов (например, гормонов); ограничивает внутреннее содержимое клетки; регулирует обмен веществ между клеткой и окружающей средой, обеспечивая постоянство внутриклеточного состава; участвует в формировании межклеточных контактов.
Рис. 12. Схема строения эукариотических клеток. Цитоплазма представляет собой внутреннее содержимое клетки и состоит из гиалоплазмы и находящихся в нем разнообразных внутриклеточных структур. Гиалоплазма (матрикс) – это водный раствор неорганических и органических веществ, способный изменять свою вязкость и находящиеся в постоянном движении. Цитоплазматические структуры клетки представлены включениями и органоидами. Включения – относительно непостоянные, встречающиеся в клетках некоторых типов в определенные моменты жизнедеятельности, например, в качестве запаса питательных веществ (зерна крахмала, белков, капли гликогена) или продуктов подлежащих выделению из клетки. Органоиды – постоянные и обязательные компоненты большинства клеток, имеющим специфическую структуру и выполняющим жизненно важную функцию. Эндоплазматическая сеть. Вся внутренняя зона цитоплазмы заполнена многочисленными мелкими каналами и полостями, стенки которых представляют собой мембраны, сходные по своей структуре с плазматической мембраной. Эти каналы ветвятся, соединяются друг с другом и образуют сеть, получившую название эндоплазматической сети. Эндоплазматическая сеть неоднородна по своему строению. Известны два ее типа - гранулярная и гладкая. На мембранах каналов и полостей гранулярной сети располагается множество мелких округлых телец - рибосом, которые придают мембранам шероховатый вид. Мембраны гладкой эндоплазматической сети не несут рибосом на своей поверхности. Основная функция гранулярной эндоплазматической сети - участие в синтезе белка, который осуществляется в рибосомах. На мембранах гладкой эндоплазматической сети происходит синтез липидов и углеводов. Эндоплазматическая сеть связывает между собой основные органоиды клетки. загрузка... Аппарат Гольджи. В состав аппарата Гольджи входят полости, ограниченные мембранами и расположенные группами (по 5-10) и крупные и мелкие пузырьки, расположенные на концах полостей. Митохондрии. Оболочка митохондрии состоит из двух мембран - наружной и внутренней. Наружная мембрана гладкая, она не образует никаких складок и выростов. Внутренняя Пластиды. В цитоплазме клеток всех растений находятся пластиды. В клетках животных пластиды отсутствуют. Различают три основных типа пластид: зеленые - хлоропласты; красные, оранжевые и желтые - хромопласты; бесцветные - лейкопласты. В пластидах, как и митохондриях, имеется две мембраны: наружная (гладкая) и внутренняя, образующая выпячивания (ламеллы и тилакоиды). В хлоропластах имеется хлорофилл, основная их функция – фотосинтез. Лизосомы - небольшие округлые тельца. Внутри лизосомы находятся ферменты, расщепляющие белки, жиры, углеводы, нуклеиновые кислоты. Обладая способностью к активному перевариванию пищевых веществ, лизосомы участвуют в удалении отмирающих в процессе жизнедеятельности частей клеток, целых клеток и органов. Вакуоль – органелла, отграниченная от цитоплазмы мембраной – тонопластом. В животных клетках могут наблюдаться небольшие вакуоли, выполняющие фагоцитарную, пищеварительную, сократительную и другие функции. Растительные клетки имеют одну большую центральную вакуоль. Жидкость, заполняющая её, называется клеточным соком. Это концентрированный раствор сахаров, минеральных солей, органических кислот, пигментов и других веществ. Вакуоли накапливают воду, могут содержать красящие пигменты, защитные вещества, ферменты, запасные питательные вещества. Обязательными для большинства клеток являются также органоиды, не имеющие мембранного строения. К ним относятся рибосомы, микрофиламенты, микротрубочки, клеточный центр. Рибосомы обнаружены в клетках всех организмов. Это Микротрубочки и микрофиламенты – нитевидные структуры, состоящие из различных сократительных белков и обуславливающие двигательные функции клетки. Микротрубочки имеют вид полых цилиндров, стенки которых состоят из белков – тубулинов. Микрофиламенты представляют собой очень тонкие, длинные, нитевидные структуры, состоящие из актина и миозина. Микротрубочки и микрофиламенты пронизывают всю цитоплазму клетки, формируя её цитоскелет. Клеточный центр. Основную часть клеточного центра составляют два маленьких тельца - центриоли, расположенные в небольшом участке уплотненной цитоплазмы. Каждая центриоль имеет форму цилиндра длиной до 1 мкм. Центриоли играют важную Ядро – наиболее важный компонент эукариотических клеток. Хроматин образован молекулами ДНК в комплексе с белками. В процессе деления клеток происходит спирализация ДНК и хроматиновые структуры образуют хромосомы. Обычно в ядрах клеток тела (соматических) хромосомы представлены парами, в половых клетках они не парны. Одинарный набор хромосом в половых клетках называют гаплоидным (n), двойной набор хромосом в соматических клетках - диплоидным (2n). Диплоидный набор хромосом клеток конкретного вида живых организмов, характеризующийся числом, величиной и формой хромосом, называют кариотипом. Несмотря на единый принцип строения, между клетками эукариотических организмов разных царств имеются различия (табл. 2).
Сравнительная характеристика эукариотических клеток
Таблица 2
Рост и размножение организмов связаны с делением клеток. Существует два основных способа деления клеток. Митоз — это такое деление клеточного ядра, при котором образуются два дочерних ядра с наборами хромосом, идентичными наборам родительской клетки. Например, за счет процессов митоза у человека постоянно возобновляется слущивающийся эпителий кожи. Мейоз — это деление клеточного ядра с образованием четырех дочерних ядер, каждое из которых содержит вдвое меньше хромосом, чем исходное ядро. За счет мейотического деления из диплоидных клеток образуются гаплоидные половые клетки, которые при слиянии (оплодотворении) восстанавливают диплоидный набор хромосом в одноклеточном зародыше – зиготе.
35. Криптохромы Ряд процессов в растительном организме контролируется исключительно синим светом (диапазон длин волн 360-480 нм): фототропизм, синтез хлорофилла и каротиноидов, фототаксис хлоропластов и др. Фоторецепторы синего света называются криптохромами. Они имеются в клетках всех тканей, локализованы в плазмалемме. Функции хромофоров выполняют флавин (ФАД, ФМН), изоаллоксазиновое кольцо содержит конъюгированную систему связей, воспринимающую энергию квантов света. Синий свет стимулирует деление, но задерживает растяжение клеток, поэтому для растений высокогорий характерно явление нанизма, т.е. низкорослости. Действие фоторецепторов проявляется через непосредственное действие на генетический аппарат растений и регуляцию экспрессии генов, в том числе тех генов, которые кодируют ферменты синтеза гормонов. А гормоны, регулирующие разные этапы метаболизма, определяют процессы роста и развития растений. Основным критерием фотопериодической реакции растений считается переход к генеративной фазе. Но как оказалось, от соотношения освещенного и неосвещенного периода суток зависят и другие явления в жизни растений: продуктивность, устойчивость к инфекционным заболеваниям, способность к симбиозам. Особо важную роль играет фотопериод в географическом распространении растений. Существует достаточно четкая связь между географическим распространением растений и типом их фотопериодической реакции. Виды, распространенные в высоких широтах, в основном относятся к длиннодневным, виды тропиков и субтропиков – к короткодневным и нейтральным. У видов с широким ареалом распространения различаются географические популяции с разными фотопериодами. У культурных растений фотопериодическая реакция (ФПР) соответствует географическому району формирования сорта. Абиссинские сорта пшеницы имеют короткодневный тип ФПР, а сибирские – длиннодневный тип ФПР. Являясь адаптацией к данной географической среде, ФПР является ограничителем распространения вида или географической популяции. Длина дня препятствует миграции северных длиннодневных форм к югу, а южных короткодневных – к северу. Виды с нейтральной ФПР имеют большую возможность для распространения. Закономерности распределения растений с разными типами ФПР не абсолютны. В горах низких широт встречаются длиннодневные растения (южноамериканские дикие пасленовые). Культурные растения, происходящие из Индии, Эфиопии (ячмень, овес, пшеница), имеют ФПР длиннодневного типа. Объяснение этому следующее: ФПР в большей степени зависит от температурного фона, в результате длиннодневные растения могут произрастать при более коротком дне при условии пониженных температур. Поэтому в горных областях близ экватора, где холоднее, могут произрастать длиннодневные растения. Возможно, что горные массивы меридионального направления могут служить своеобразными мостами для миграции длиннодневных растений из северных широт в более южные. Вторым важным фактором, влияющим на переход к генеративной фазе, являются низкие положительные температуры. Индуцирование процесса цветения низкими температурами называется яровизацией. Все растения делят на 3 группы: 1. Растения, которые не переходят к генеративной фазе без воздействия низких температур (озимые зерновые, двулетние растения – морковь, свекла, капуста и некоторые многолетники). 2. Растения, у которых развитие ускоряется под воздействием пониженных температур. 3. Растения, которые не требуют для перехода к цветению пониженных температур. Растения, которые не нуждаются в воздействии низких температур, называют яровыми, нуждающиеся – озимыми. Низкие температуры необходимы для формирования флорального стимула. Для яровизации озимых форм, с тем чтобы их можно было высевать весной, необходимым условием являются: наличие делящихся клеток и воздействие температур 1-5оС в течение 1-3 месяцев. Для растений южных широт этот температурный диапазон около 10оС. Причем некоторые растения (злаки) могут воспринять яровизационные воздействия в стадии наклюнувшихся семян, другие же (капуста, сельдерей) - только при наличии листьев. Выявлено, что в результате яровизации повышается концентрация гиббереллина, а он, как известно, является компонентом флоригена. Яровизационные изменения обратимы, под действием высоких температур возможна разъяровизация. 36. Светолюбивые растения - гелиофиты, растения, произрастающие на открытых местах и не выносящие длительного затенения; для нормального роста им необходима интенсивная солнечная или искусственная радиация. Взрослые гелиофиты, как правило, более светолюбивы, чем молодые экземпляры. К светолюбивым растениям относятся как травянистые (подорожник большой, кувшинка и др.), так и древесные (лиственница, акация и др.) растения, ранневесенние - степей и полупустынь, а из культурных — кукуруза, сорго, сахарный тростник и др. Древесные или кустарниковые гелиофиты образуют обычно разреженные посадки. Листья – равносторонние, узкие блестящие; побеги короткие; есть волоски. Приспособленность к интенсивному освещению обеспечивается особенностями морфологии и физиологии светолюбивых растений. У них обычно довольно толсты
|
|||||||||||||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-08-01; просмотров: 448; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.216.161.178 (0.016 с.) |