Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Влияние внешних и внутренних факторов на фотосинтезСодержание книги
Поиск на нашем сайте Организм человека, Физиологическая функция – это проявление взаимодействия между отдельными частями, элементами структуры живой системы. В физиологических функциях проявляется жизнедеятельность как целостного организма, так и отдельных его частей. Физиологические процессы и физиологические функции изучаются в организме в целом, в его системах, органах, тканях и клетках. Организм – это открытая, самостоятельно существующая, саморегулирующаяся единица органического мира, которая реагирует как единое целое на изменение условий окружающей среды. Организм можно изучать на системном, органном, тканевом, клеточном и молекуляном уровнях. Клетка – элементарная живая система, основная структурная и функциональная единица организма. Она способна к самообновлению, саморегуляции и самовоспроизведению. Клетки объединяются в ткани. Ткань – представляет собой совокупность клеток и межклеточного вещества, имеющих одинаковое происхождение, строение и выполняющих в организме определенную деятельность (например, мышечная ткань осуществляет сократительную деятельность). Ткани образуют органы. Орган – это часть тела, имеющая определенную форму, состоящая из различных тканей и структур, которые объединены для выполнения специфического вида деятельности (например, почки – орган образования мочи). Органы объединены в системы. Система органов – это объединение органов, совместно выполняющих общие функции.
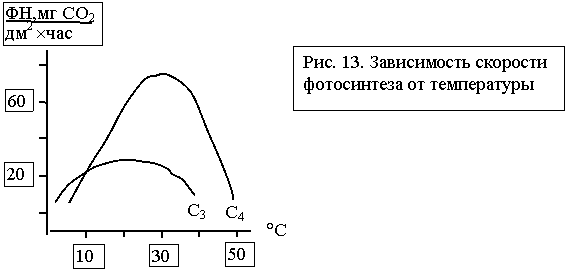
Органы и ткани состоят из скопления клеток, размеры, формы и число которых различны в зависимости от органа и выполняемой им функции. Животная клетка – это структурно-функциональная единица живого организма, способная к делению и обмену с окружающей средой. Она осуществляет передачу генетической информации путем самовоспроизведения. Деятельностью клеток определяется деятельность всех тканей и органов. Существует несколько типов клеток - эпителиальные - мышечные - нервные - клетки крови - костные - клетки соединительной ткани. Каждая клетка имеет сложное строение и представляет собой систему биополимеров, содержит ядро, цитоплазму и находящиеся в ней органеллы. От внешней среды клетка отграничивается клеточной оболочкой – плазмалеммой, которая осуществляет транспорт необходимых веществ в клетку, и наоборот, взаимодействует с соседними клетками и межклеточным веществом. Внутри клетки находится ядро, в котором происходит синтез белка, оно хранит генетическую информацию в виде ДНК. Ядро может иметь округлую или овоидную форму, но в плоских клетках оно несколько сплющенное, а в лейкоцитах палочковидное или бобовидное. В эритроцитах и тромбоцитах оно отсутствует. Сверху ядро покрыто ядерной оболочкой, которая представлена внешней и внутренней мембраной. В ядре находится нуклеоплазма, которая представляет собой гелеобразное вещество и содержит хроматин и ядрышко. Ядро окружает цитоплазма, в состав которой входит гиалоплазма, органеллы и включения. Гиалоплазма – это основное вещество цитоплазмы, она участвует в обменных процессах клетки, содержит белки, полисахариды, нуклеиновую кислоту и др. Постоянные части клетки, которые имеют определенную структуру и выполняют биохимические функции, называют органеллами. К ним относятся клеточный центр, митохондрии, комплекс Гольджи, эндоплазматическая (цитоплазматическая) сеть. Клеточный центр обычно находится около ядра или комплекса Гольджи, состоит из двух плотных образований – центриолей, которые входят в состав веретена делящейся клетки и образуют реснички и жгутики. Митохондрии имеют форму зерен, нитей, палочек, формируются из двух мембран – внутренней и внешней. Внутренняя мембрана образует складки (кристы), в которых располагаются ферменты. В митохондриях происходят расщепление глюкозы, аминокислот, окисление жирных кислот, образование АТФ – основного энергетического материала. Комплекс Гольджи (внутриклеточный сетчатый аппарат) имеет вид пузырьков, пластинок, трубочек, расположенных вокруг ядра. Его функция состоит в транспорте веществ, химической их обработке и выведении за пределы клетки продуктов ее жизнедеятельности. Эндоплазматический (цитоплазматический) ретикулум или сеть формируется из агранулярной (гладкой) и гранулярной (зернистой) сети. Агранулярная эндоплазматическая сеть образуется преимущественно мелкими цистернами и трубочками, которые участвуют в обмене липидов и полисахаридов. Гранулярная эндоплазматическая сеть состоит из пластинок, трубочек, цистерн, к стенкам которых прилегают мелкие образования – рибосомы, синтезирующие белки. Цитоплазма также имеет постоянные скопления отдельных веществ, которые называются включениями цитоплазмы и имеют белковую, жировую и пигментную природу. Клетка как часть многоклеточного организма выполняет основные функции: усвоение поступающих веществ и расщепление их с образованием энергии, необходимой для поддержания жизнедеятельности организма. Клетки обладают также раздражимостью (двигательные реакции) и способны размножаться делением. Деление клеток бывает непрямое (митоз) и редукционное (мейоз). загрузка... Итак, как мы уже говорили, клетка входит в состав ткани, из которой состоит организм человека и животных. В результате взаимодействия организма с внешней средой, которое сложилось в процессе эволюции, появились четыре вида тканей с определенными функциональными особенностями: эпителиальная, соединительная, мышечная и нервная. Каждый орган состоит из различных тканей, которые тесно связаны между собой. Например, желудок, кишечник, другие органы состоят из эпителиальной, соединительной, гладкомышечной нервной тканей. Соединительная ткань многих органов образует строму, а эпителиальная – паренхиму. Таким образом, различные ткани, входящие в состав того или иного органа, обеспечивают выполнение главной функции данного органа. Как мы уже говорили, все клетки объединяются в ткани, ткани в органы, а органы в системы органов. 5. Процесс обмена азотистых веществ происходит в течение всей жизни растения, но характер и темпы этого процесса в разные периоды роста и развития неодинаковы. Температура Температура оказывает влияние на ход всех ферментных реакций, поэтому отчетливо сказывается на скорости фотосинтеза (рис. 13). У некоторых видов хвойных деревьев процесс фотосинтеза протекает и при отрицательных температурах, но у всех полевых культур прекращается при нескольких градусах выше нуля. На рисунке 13 также заметно существенное преимущество растений С4 с точки зрения активности фотосинтеза при повышении температур. И наоборот, у растений С3 при одинаковых прочих условиях скорость фотосинтеза возрастает при более низких температурах. Следовательно, можно сделать практический вывод: для повышения скорости фотосинтеза растениям С4 необходимы благоприятные температурные условия.
Однако работа с кукурузой показывает, что варьирующую зависимость скорости фотосинтеза от температуры можно использовать путем целенаправленной селекции при получении высокопродуктивных сортов для относительно холодных условий. Следует отметить, что успешное возделывание кукурузы в холодных областях не обусловлено лишь взаимосвязью между скоростью фотосинтеза и температурой, хотя именно эту зависимость используют в селекции данной культуры. Влияние воды Влияние воды на скорость фотосинтеза выражено величиной дефицита насыщения, отражающей то количество воды, которого не хватает растению для полного насыщения. Максимальной скорости фотосинтеза растения достигают при очень низких показателях дефицита: при 12-20% устьица заполнены водой с скорость фотосинтеза падает до нуля (подобные сведения в литературе [166,190,195]). Основной предпосылкой для фотосинтеза является наличие хлорофилла. Его количество колеблется в пределах 2-6 мг на 0,01 м2 площади листьев, что примерно соответствует 0,5-2,0% сухого вещества листа. Пока в исследованиях не удалось установить зависимости скорости фотосинтеза от содержания хлорофилла. Следовательно, у здоровых растений, развивающихся в нормальных условиях, недостаток хлорофилла не отражается на ФН. Он становится лимитирующим фактором лишь для листьев, растущих при недостатке света, дефиците воды и минеральных веществ или при поражении некоторыми болезнями. Минеральные вещества Научные исследования и практика показывают, что на фотосинтез оказывают существенное влияние минеральные вещества. Дефицит любого из следующих элементов –N, P, K, Mg, S, Ca, Fe, Mn, Cu, B, Zn, Mo, - отрицательно сказывается на скорости фотосинтеза. Воздействие питательных веществ на фотосинтетический аппарат бывает многосторонним. Они влияют на формирование морфологической и анатомической структуры растения, на физические процессы, участвуют в создании хлоропластов, причем входят в состав как отдельных структурных образования, так и ферментов. Определение количественных характеристик оптимальных условий минерального питания – довольно трудная задача. Определенное исключение составляет лишь азот. У многих видов растений установлена зависимость между содержанием азота в ассимилирующих тканях и скоростью фотосинтеза. Например, показанная на рисунке 14 зависимость ФН от количества азота в пластинках верхних листьев озимой пшеницы в полевых условиях. На рисунке также показаны величины обычного содержания азота в листьях.
Возраст Возраст отдельных органов и растения в целом является существенным эндогенным фактором, определяющим скорость фотосинтеза. У молодого листа ФН увеличивается до определенного максимума, достигая его еще перед окончанием роста листа, т.е. до формирования его полной площади. Этот период называется фотосинтетической зрелостью листа и длится разное время.
7. Диффузия воды через полупроницаемую мембрану называется осмосом. Полупроницаемая мембрана – это мембрана, хорошо проницаемая для воды и непроницаемая или плохо проницаемая для растворенных в воде веществ. Осмотическая ячейка – это пространство, окруженное полупроницаемой мембраной и заполненное каким-либо водным раствором, способным развивать определенное осмотическое давление. Осмотическое давление (диффузное давление) – термодинамический параметр, характеризующий стремление раствора к понижению концентрации при соприкосновении с чистым растворителем вследствие встречной диффузии молекул растворённого вещества и растворителя. Если раствор отделен от чистого растворителя полупроницаемой мембраной, то возможна лишь односторонняя диффузия – осмотическое всасывание растворителя через мембрану в раствор. В этом случае осмотическое давление становится доступной для прямого измерения величиной. Оно равно избыточному давлению, приложенному со стороны раствора при осмотическом равновесии. Осмотическое давление обусловлено понижением химического потенциала растворителя в присутствии растворённого вещества. Тенденция системы выравнивать химические потенциалы во всех частях своего объёма и переходить в состояние с более низким уровнем свободной энергии вызывает осмотический (диффузионный) перенос вещества. Осмотическое давление в идеальных и предельно разбавленных растворах не зависит от природы растворителя и растворённых веществ; при постоянной температуре оно определяется только числом «кинетических элементов» (ионов, молекул, ассоциатов или коллоидных частиц) в единице объёма раствора. Осмотическое давление (Р) численно равно давлению, которое оказало бы растворённое веще- ство, если бы оно при данной температуре находилось в состоянии идеального газа и занимало объём, равный объёму раствора. Осмотическое давление измеряют с помощью специальных приборов (осмометров), определяя избыточное гидростатическое давление столба жидкости в трубке осмометра после установления осмотического равновесия. Осмос является основным механизмом поступления воды в растительную клетку. Все клеточные мембраны, в том числе плазмалемма и тонопласт, являются полупроницаемыми мембранами. Вода проходит в клетку через водные поры в плазмалемме, образованные специальными белками – аквапоринами. Внутри вакуоли («осмотической ячейки») клеточный сок развивает ос- мотическое давление π: π = i С R T, где С – концентрация раствора в молях; Т – абсолютная температура; R – газовая постоянная 0,082 л × атм/град × моль; i – изотонический коэффициент, равный 1 + α(n–1), где α – степень электролитической диссоциации; n – число ионов, на которые распадается молекула электролита.
Благодаря осмотическому притоку воды в клетку там возникает гидростатическое давление, называемое тургорным. Это давление прижимает цитоплазму к клеточной стенке и растягивает ее. Клеточная стенка имеет ограниченную эластичность и оказывает равное противодавление. Эластическое растяжение ткани благодаря тургорному давлению ее клеток придает твердость неодревесневшим частям растений. Завядающие побеги становятся дряблыми, так как при потере воды тургорное давление падает. Тургорное давление противодействует притоку воды в клетку. Давление, с которым вода осмотически притекает в клетку, равно, таким образом, разности осмотического давления π и тургорного давления P. Эту величину называют сосущей силой S: S = π – P. Вода поступает в клетку из внешнего раствора, если его потенциальное осмотическое давление меньше сосущей силы клетки, и, наоборот, вода выходит из клетки в раствор с более высоким потенциальным осмотическим давлением. Величину осмотического потенциала можно определить плазмолитическим методом. Плазмолиз – это процесс, обусловленный потерей воды клеткой. Он проявляется в отходе протопласта от клеточной стенки. При переносе плазмолизированных тканей в гипотонический раствор (или чистую воду) вода поступает в клетку и происходит деплазмолиз. Количество воды в клетке увеличивается, объем вакуоли возрастает – и она прижимает цитоплазму к клеточной стенке. Плазмолитический метод основан на подборе изоосмотического (изотонического) раствора, то есть имеющего осмотический потенциал, равный осмотическому потенциалу клетки. Раствор, при котором начался плазмолиз, имеет осмотический потенциал, примерно равный осмотическому потенциалу клетки. Зная концентрацию наружного раствора в молях, можно вычислить осмотический потенциал клетки. Иногда при сильном завядании протопласт не отстает от клеточной стенки, как при плазмолизе, а сжимается и тянет ее за собой. При этом клеточная стенка прогибается. Это явление называют циторризом. Развивается натяжение (или отрицательное давление стенки) – и потенциал тургорного давления приобретает отрицательное значение. В этом случае величина водного потенциала определяется уже не разностью, а суммой осмотического загрузка...
Величина осмотического потенциала позволяет судить о способности растения поглощать воду из почвы и удерживать ее, несмотря на иссушающее действие атмосферы. Осмотический потенциал колеблется у разных растений в пределах от –5 до –200 баров. У водных растений осмотический потенциал около –1 бара. У большинства растений средней полосы осмотический потенциал колеблется от –5 до –30 баров, растения степей и пустынь имеют более отрицательный осмотический потенциал. Осмотический потенциал различен и у разных жизненных форм. У деревьев он отрицательнее, чем у кустарников и травянистых растений. У светолюбивых растений осмотический потенциал отрицательнее, чем у теневыносливых растений. Поступление воды в клетку обусловлено не только осмотическим давлением, но и силой набухания. Набуханием называют поглощение жидкости или пара высокомолекулярным веществом (набухающим телом), сопровождаемое увеличением объема. Явление набухания обусловлено коллоидальными и капиллярными эффектами. В протоплазме преобладает набухание на коллоидальной основе (гидратация коллоидов), а в клеточной стенке наблю-даются оба эффекта: капиллярный – накопление воды между микрофибриллами и в межмицеллярных пространствах; коллоидальный – гидратация полисахаридов, особенно гемицеллюлоз. У некоторых частей растений поглощение воды происходит исключительно путем набухания, например у семян.
Вода в клетку может поступать также в процессе пиноцитоза, когда часть плазмалеммы прогибается внутрь клетки. Внешние края такой инвагинации смыкаются и виде пузырька – везикулы с адсорбированными частицами и внешним раствором, – который проходит внутрь цитоплазмы. 8. Ростовые корреляции (стимулирующие, ингибирующие, компенсаторные) - отражают зависимость роста и развития одних органов или частей растения от других, их взаимное влияние. Примером стимулирующих корреляций является взаимовлияние побега и корня. Корень обеспечивает надземные органы водой, питательными веществами, а из листьев в корни поступают органические вещества (углеводы, ауксины), необходимые для роста корней. Ингибирующие корреляции (тормозящие) - о дни органы подавляют рост и развитие других органов. Примером данных корреляций может служить явление а пикального доминирования – торможение роста боковых почек, побегов верхушечной почкой побега. Примером может служить феномен «королевского» плода, завязавшегося первым. Использование в практике приема снятия апикального доминирования: формирование кроны путем обрезки верхушек доминирующих побегов, пикировка рассады и саженцев плодовых деревьев. Компенсаторные корреляции отражают зависимость роста и конкурентные отношения отдельных органов от обеспеченности их питательными в-вами. В процессе роста растительного организма идет естественная редукция (опадение, отмирание) или искусственно удаляется часть развивающихся органов (пасынкование, прореживание завязей), а оставшиеся растут с большей скоростью. Регенерация - восстановление поврежденных или утраченных частей.
Полярность - свойственная растениям специфическая дифференциация структур и процессов в пространстве. Она проявляется в определенной направленности роста корня и стебля, в определенном направлении передвижения веществ. 9. Физиология растений — наука, которая изучает процессы жизнедеятельности и функции растительного организма. Слово «физиология» греческого происхождения; оно состоит из двух слов: physis — природа и logos — понятие, учение. Физиология растений является наиболее развитой отраслью экспериментальной ботаники, которая в XIX в. выделилась в самостоятельную науку. Она тесно связана с химией, физикой, биохимией, биофизикой, микробиологией, молекулярной биологией. Перед научными работниками, физиологами растений поставлены такие задачи: изучить обмен веществ и энергии в растительном организме, фотосинтез, хемосинтез, биологическую фиксацию азота из атмосферы и корневое питание растений; разработать методы повышения использования растениями солнечной энергии и питательных веществ почвы, обогащения почвы азотом; создать новые, более эффективные формы удобрений и разработать методы их применения; исследовать действие биологически активных веществ с целью использования их в растениеводстве; разработать методы более продуктивного использования воды растением. Без решения этих вопросов невозможно решение и ряда других проблем земледелия и растениеводства, направленных на повышение урожайности. Интенсивное применение минеральных удобрений, гербицидов, физиологически активных веществ, химических препаратов для защиты растений от болезней и вредителей требует глубокого и всестороннего изучения их влияния на рост и обмен веществ растительных организмов с целью значительного повышения продуктивности сельскохозяйственных растений. Решение поставленных задач имеет большое значение для разработки проблем ускорения научно-технического прогресса в растениеводстве и дальнейшего развития сельского хозяйства нашей страны. Основной метод познания процессов, явлений в физиологии — эксперимент, опыт. Следовательно, физиология растений — наука экспериментальная. Для изучений физико-химической сути функций, процессов в физиологии растений широко применяют методы: лабораторно-аналитический, вегетационный, полевой, меченых атомов, электронной микроскопии, электрофореза, хроматографического анализа, ультрафиолетовой и люминесцентной микроскопии, спектрофотометрии и др. Кроме того, используют фитотроны и лаборатории искусственного климата, в которых выращивают растения и проводят опыты в условиях определенного состава воздуха, нужной температуры и освещения. Применяя эти методы, физиологи исследуют растения на молекулярном, субклеточном, клеточном и организменном (интактное растение) уровнях. Сейчас в биологических исследованиях широко применяют электронные микроскопы просвечивающего типа с разрешающей способностью 0,15—0,5 нм, в которых объект рассматривают в электронных лучах, проходящих через него. Значительное увеличение разрешающей способности электронных микроскопов по сравнению со световыми обусловливается меньшей длиной волны электронов (на пять порядков меньшей, чем длина волны ультрафиолетовых лучей). Кроме того, для биологических исследований применяют так называемые растровые электронные микроскопы, в которых изображение создается по принципу телевизионных. Разрешающая способность растровых микроскопов равна 20—40 нм, с их помощью изучают строение поверхности пыльцы, эпидермального слоя клеток, формы клеток и др. Применение электронной микроскопии в биологии имеет большое значение для развития биологической науки и физиологии растений в частности. Исследование ультраструктуры органоидов растительной клетки (хлоропластов, митохондрий, рибосом, мембранных структур) дало возможность раскрыть суть процессов фотосинтеза и дыхания, которые определяют возможность самой жизни на нашей планете. Изучение строения клеточных оболочек, открытие цитоплазматических мембранных структур способствовали выяснению процессов обмена веществ и энергии в клетке, изучению структуры и функции органоидов растительной клетки. Большое принципиальное значение имеет электронно-микроскопическое исследование строения РНК и ДНК, локализации их на структурных компонентах клетки. Результаты этих исследований легли в основу раскрытия генетической роли ядра и проблемы наследственности. Процесс нитрификации Продукты гниения белков и разложения мочевины — аммиак и аммиачные соли — могут быть непосредственно усвоены растениями, но они обычно превращаются в нитраты — соли азотной кислоты. Биологическая сущность процесса нитрификации была доказана работами Т. Шлезинга и Л. Мюнца в 1879 г. Позднее (1888 — 1890) известный русский микробиолог С. Н. Виноградский, применив элективную среду, выделил чистые культуры тарификаторов. Ученый установил, что органическое вещество в среде тормозит развитие нитрифицирующих бактерий, в то время как в чисто минеральных питательных растворах они хорошо растут. В первой фазе аммиак окисляется до азотистой кислоты по схеме: NH3 → NH4OH → NH2OH → HNO → HNO +274,9 кДж. Считается, что процесс нитрификации проходит в несколько стадий, при этом образуется ряд промежуточных продуктов: гидроксиламин, нитроксил и др. Во второй фазе азотистая кислота окисляется до азотной: HNO2 → HNO3 +87,6 кДж. Первая и вторая фазы единого процесса нитрификации вызываются разными возбудителями. С. Н. Виноградский объединил их в три рода; Nitrosomonas, Nitrococystis, Nitrosospira. Бактерии рода Nitrosomonas имеют форму палочек, грамотрицательные, подвижные, снабжены одним жгутиком, спор не образуют. Разные виды Nitrosomonas широко распространены в почве и отличаются друг от друга формой и размерами. Род Nilrococystis способен образовывать зооглеи (кокковые формы микробов окружены общей капсулой). Род Nitrosospira С. Н. Виноградский разделил на два вида: Nitrosospira bria и Nitrosospira arctica. Бактерии обоих видов имеют правильную спиральную форму. Наряду со спирально закрученными нитями у старых культур встречаются короткие палочки и кокки. В последнее время выделено еще два рода микробов, вызывающих первую фазу нитрификации: Nitrosolobus и Nitrosovibrio. Окисление азотистой кислоты в азотную осуществляется мелкой полиморфной, грамотрицательной, неподвижной бактерией, которую С. Н. Виноградский назвал Nitrobacter, Микробы группы Nitrobacter лучше развиваются на чисто минеральных средах и могут синтезировать органическое вещество своего тела, используя углерода диоксид. Во второй фазе нитрификации применяют участие также микробы родов Nitrospina и Nitrococcus. Нитрифицирующие микробы отрицательно относятся к органическим веществам. Добавление к минеральному раствору 0,2% пептона или глюкозы приостанавливает рост микробов. Сильная чувствительность нитрифицирующих микробов к органическим веществам отмечается в растворах, в почве этого не наблюдается, так как в ней водорастворимых веществ в значительных количествах никогда не бывает. Кроме нитрифицирующих бактерий, в почве находятся и другие микроорганизмы, которые используют органическое вещество и тем самым создают благоприятные условия для развития нитрификаторов. Это лишний раз свидетельствует о том, что физиологические свойства микроорганизмов надо изучать не на изолированных искусственных средах, а в естественной среде их обитания. На процессы окисления аммиака влияют не только микробы, но и их ферменты. Кроме органического вещества, на нитрификацию оказывает влияние концентрация аммиака. Его действие на культуру резко проявляется в условиях жидких сред. В почве же аммиак, находится в адсорбированном состоянии и не может оказывать угнетающего действия. Поэтому нитробактер сразу же окисляет азотистую кислоту в азотную. На процесс нитрификации положительно сказывается присутствие кислорода. В обрабатываемых почвах процесс нитрификации протекает более интенсивно. Примерно также нитрификация проходит на черноземных почвах, особенно когда в них достаточное количество аммонифицирующих микробов, готовящих пищу (среду) для нитрификаторов. У солонцов меньшая нитрифицирующая способность. В почвах азотистая кислота не накапливается, поскольку Nitrosomonas и Nitrobacter встречаются в одной среде, находятся в своеобразном симбиозе. Нитрификаторы способны осуществлять хемосинтез, то есть создавать органическое вещество из углерода диоксида и воды за счет химической энергии окисления аммиака до азотистой кислоты и азотистой до азотной кислоты. Нитрификаторы чувствительны к кислой среде, они лучше развиваются при рН 8,3—9,3. В результате жизнедеятельности нитрифицирующих бактерий на 1 га почвы может накапливаться за год до 300 кг азотной кислоты. Д.Н. Прянишников установил, что одним из важнейших условий использования аммонийных и нитратных солей является реакция среды: в слабокислой среде при рН 5 лучше усваиваются нитраты, и, наоборот, в нейтральной среде при рН5лучше поглощаются аммонийные соли. Для использования аммонийных солей необходимо достаточное количество углеводов в растениях, без которых задерживается превращение их в амиды, накапливается аммиак, действующий токсический на растение. При питании растений физиологический нейтральной солью (NН4NO3) и реакции среды, близкой к нейтральной, лучше усваивается катион NН4+, чем анион NО3-, в этом случае соль будет физиологический кислой. Таким образом, реакция среды имеет большое значение для усвоения растениями соединений, содержащих азот. Экспериментально доказано, что преимущественное поглощение аммонийных солей свойственно растениям, склонным к усиленному образованию органических кислот.
12. Химический состав растений, как известно, отражает элементный состав почв. Поэтому избыточное накопление ТМ растениями обусловлено, прежде всего, их высокими концентрациями в почвах. В своей жизнедеятельности растения контактируют только с доступными формами ТМ, количество которых, в свою очередь, тесно связано с буферностью почв. Однако способность почв связывать и инактивировать ТМ имеет свои пределы, и когда они уже не справляются с поступающим потоком металлов, важное значение приобретает наличие у самих растений физиолого-биохимических механизмов, препятствующих их поступлению. Механизмы устойчивости растений к избытку ТМ могут проявляться по разным направлениям: одни виды способны накапливать высокие концентрации ТМ, но проявлять к ним толерантность; другие стремятся снизить их поступления путем максимального использования своих барьерных функций. Для большинства растений первым барьерным уровнем являются корни, где задерживается наибольшее количества ТМ, следующий – стебли и листья, и наконец, последний ≈ органы и части растений, отвечающие за воспроизводительные функции. Уровень накопления ТМ разными растениями в зависимости от их генетических и видовых особенностей при одинаковом содержании ТМ в почвах. Отмечаются случаи, когда разные сорта одной культуры, произрастающие на одинаково загрязненной почве, содержали различные количество ТМ. Данный факт, по-видимому, обусловлен присущим всем живым организмам внутривидовым полиморфизмом, способным проявить себя и при техногенном загрязнении природной среды. Это свойство у растения может стать основой генетически – селекционных исследований с целью создания сортов с повышенными защитными возможностями по отношению к избыточным концентрациям ТМ. Несмотря на существенную изменчивость различных растений к накоплению ТМ, биоаккумуляция элементов имеет определенную тенденцию, позволяющую упорядочить их в несколько групп: 1) Cd, Cs, Rb – элементы интенсивного поглощения; 2) Zn, Mo, Cu, Pb, As – средней степени поглощения; 3) Mn, Ni – слабого поглощения; 4) Se, Fe, Ва – элементы, труднодоступные растениям. Другой путь поступления ТМ в растения ≈ некорневое поглощение из воздушных потоков. Оно имеет место при значительном выпадении металлов из атмосферы на листовой аппарат, чаще всего вблизи крупных промышленных предприятий. Поступление элементов в растения через листья (или фолиарное поглощение) происходит, главным образом, путем неметаболического проникновения через кутикулу. ТМ, поглощенные листьями, могут переноситься в другие органы и ткани и включаться в обмен веществ. Токсичность тяжелых металлов связана с их физико-химическими свойствами, со способностью к образованию прочных соединений с рядом функциональных группировок на поверхности и внутри клеток. Таким образом, действие металлов на растительный организм зависит от природы элемента, содержание его в окружающей среде, характера почвы, формы химического соединения, срока от момента загрязнения. Формирование химического состава растительного организма определяется биохимическими особенностями различных видов организмов, их возрастом и биохимическими закономерностями связи между элементами в организме. Содержание одних и тех химических элементов в различных частях растений. Важную роль в защите растений от избытка поступающих из почвы в корни ТМ выполняет корневая система. Задерживая избыточные ионы, корни тем самым способствуют сохранению в наземных органах невредных (благоприятных) концентрации химических элементов. Связывание ТМ органическими соединениями не всегда означает перевод в малоподвижное состояние. Отмечается, что некоторые комплексы ТМ с органическими лигандами способны проходить через мембраны легче, чем ионы этих металлов, и затем внутри клетки распадаться. При сильном загрязнении среды обитания поток ТМ становится столь большим, что может наблюдаться повышенное содержание их не только в вегетативных органах, но и в органах запаса ассимилятов. Растения выглядят угнетенными (хлорозы, некрозы), снижается их продуктивность. Это указывает на нарушение течения метаболических процессов. При очень сильном загрязнении среды обитания растения прекращают развитие, гибнут. Растения загрязняются также с поверхн
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-08-01; просмотров: 711; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.108 (0.013 с.) |