Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Превращение азотистых веществ в растенияхСодержание книги
Поиск на нашем сайте
Азот входит в состав белков и нуклеиновых кислот. Белки в растении находятся в виде важных структурных компонентов цитоплазмы и ферментов. Нуклеиновые кислоты содержатся в ядре растительной клетки, цитоплазме и определяют наследственность организма. Важная роль азота связана и с тем, что он входит в состав хлорофилла, витаминов, липоидов, фосфатидов, алкалоидов, гликозидов и других органических соединений. В обмене веществ азот постоянно обновляется в составе конституционных и запасных веществ. Азот регулирует толщину клеточных стенок, продолжительность фаз формирования клеток, характер дифференциации и другие процессы. Содержание азота в растениях в среднем 3—5, а в белках 16 — 18 % сухой массы. В онтогенезе количество азота увеличивается до цветения, а после цветения уменьшается за счет потребления созревающими плодами. Максимум азота содержится в зерне, меньше — в листьях и минимум — в стеблях. Из небелковых органических соединений азота в растениях исключительное место принадлежит аминокислотам, занимающим центральное положение во всем азотном обмене растений. Небелковые органические соединения встречаются преимущественно в вегетативных органах растений, содержание их относительно выше в ранних фазах развития растения. Общее содержание небелкового органического азота в вегетативных органах растений обычно составляет не более 20— 25% общего количества азота в растении. При неблагоприятных условиях питания и недостатке калия, а также при недостаточном освещении содержание небелковых азотистых соединений значительно возрастает. Кроме органических форм азота, в растении имеются неорганические соединения его в виде нитратов, нитритов и аммиака. Особенно большие количества нитратов накапливаются в некоторых диких растениях (марь, крапива и др.). Из культурных растений наиболее богаты нитратами листья свеклы, стебли картофеля, табака, гречихи. Содержание нитратов для одних и тех же видов растений сильно изменяется в зависимости от уровня азотного питания и обеспеченности другими элементами питания; недостаток последних тормозит процесс переработки нитратов в органическую форму, и ведет к повышенному накоплению их в растении. Аммиак накапливается при резких нарушениях обмена веществ в растении в результате патологических процессов, а также при внесении аммиачных удобрений на фоне недостаточного калийного питания. Аммиак оказывает токсическое действие на растительную клетку. Нитриты, могут быть обнаружены в незначительных количествах только у растений, находящихся в условиях частичного анаэробиоза. Недостаток азота вызывает задержку роста и очень слабое развитие растений, особенно листьев и генеративных органов. Усиление азотного питания при достаточной обеспеченности растений другими элементами резко улучшает рост и развитие растений. Однако избыток азота при относительно слабом фосфатном и калийном питании растении часто приводит к отрицательным последствиям: задержке и неравномерности созревания, склонности к полеганию у злаков, большой поражаемости растений грибными и бактериальными заболеваниями, пониженной сопротивляемости растений неблагоприятным климатическим условиям. Различают несколько видов соединений азота, а именно: органические соединения – азот органический, соли аммиака – азот аммиачный и соли азотной и азотистой кислот – азот нитратный. Различные формы азота, содержащиеся в почве, - основной источник этого элемента для питания растений. Основная масса азота в почве – это органический азот; содержание аммиачного и нитратного азота невелико и на протяжении весеннего, летнего и осеннего сезонов значительно колеблется. В почве содержатся такие также органических остатки растений (корни, стебли), которые разлагаются до аммиачных и азотнокислотных солей, определенная часть азота в ней сосредотачивается в виде органических соединений. Известно, что органические вещества в почве подвергаются разложению микроорганизмами, образующие различные продукты жизнедеятельности, которые усваиваются корневой системой растений. Поэтому вопрос, усваиваются ли органические формы азота высшими зелеными растениями, можно решить лишь с помощью метода стерильных культур. Из всех соединений, содержащихся в почве, лучшими источниками азота являются аммиачные соли и соли азотной кислоты. Азот и углерод, содержащийся в навозе, органических остатках, могут использовать растением после того, как под действием бактерий они превратятся в неорганические соединения. Таким образом, существует тесная связь между питанием зеленых растений и деятельностью почвенных микроорганизмов. Процесс разложения белков, аминокислот, мочевины и других органических азотистых веществ в почве называется аммонификацией., а почвенные организмы осуществляющие этот процесс – амонификаторами. Они обладают активными ферментами, способствующему быстрому разложению белков до аминокислот, которые дезоминируются с образованием NH3. Минерализация органического азота, начатую аммонификацией, завершает процесс нитрификации, осуществляемый хемосинтезирующими нитрифицирующими бактериями – аэробами. Нитрифицирующие бактерии за счет энергии окисления могут усваивать СО2 атмосферы или карбонатов и использовать для синтеза органических веществ клетки. Интенсивность нитрификации является плодородием почвы. Корневая система растений способна непосредственно поглощать аммонийные и нитратные соли. Процесс нитрификации Продукты гниения белков и разложения мочевины — аммиак и аммиачные соли — могут быть непосредственно усвоены растениями, но они обычно превращаются в нитраты — соли азотной кислоты. Биологическая сущность процесса нитрификации была доказана работами Т. Шлезинга и Л. Мюнца в 1879 г. Позднее (1888 — 1890) известный русский микробиолог С. Н. Виноградский, применив элективную среду, выделил чистые культуры тарификаторов. Ученый установил, что органическое вещество в среде тормозит развитие нитрифицирующих бактерий, в то время как в чисто минеральных питательных растворах они хорошо растут. В первой фазе аммиак окисляется до азотистой кислоты по схеме: NH3 → NH4OH → NH2OH → HNO → HNO +274,9 кДж. Считается, что процесс нитрификации проходит в несколько стадий, при этом образуется ряд промежуточных продуктов: гидроксиламин, нитроксил и др. Во второй фазе азотистая кислота окисляется до азотной: HNO2 → HNO3 +87,6 кДж. Первая и вторая фазы единого процесса нитрификации вызываются разными возбудителями. С. Н. Виноградский объединил их в три рода; Nitrosomonas, Nitrococystis, Nitrosospira. Бактерии рода Nitrosomonas имеют форму палочек, грамотрицательные, подвижные, снабжены одним жгутиком, спор не образуют. Разные виды Nitrosomonas широко распространены в почве и отличаются друг от друга формой и размерами. Род Nilrococystis способен образовывать зооглеи (кокковые формы микробов окружены общей капсулой). Род Nitrosospira С. Н. Виноградский разделил на два вида: Nitrosospira bria и Nitrosospira arctica. Бактерии обоих видов имеют правильную спиральную форму. Наряду со спирально закрученными нитями у старых культур встречаются короткие палочки и кокки. В последнее время выделено еще два рода микробов, вызывающих первую фазу нитрификации: Nitrosolobus и Nitrosovibrio. Окисление азотистой кислоты в азотную осуществляется мелкой полиморфной, грамотрицательной, неподвижной бактерией, которую С. Н. Виноградский назвал Nitrobacter, Микробы группы Nitrobacter лучше развиваются на чисто минеральных средах и могут синтезировать органическое вещество своего тела, используя углерода диоксид. Во второй фазе нитрификации применяют участие также микробы родов Nitrospina и Nitrococcus. Нитрифицирующие микробы отрицательно относятся к органическим веществам. Добавление к минеральному раствору 0,2% пептона или глюкозы приостанавливает рост микробов. Сильная чувствительность нитрифицирующих микробов к органическим веществам отмечается в растворах, в почве этого не наблюдается, так как в ней водорастворимых веществ в значительных количествах никогда не бывает. Кроме нитрифицирующих бактерий, в почве находятся и другие микроорганизмы, которые используют органическое вещество и тем самым создают благоприятные условия для развития нитрификаторов. Это лишний раз свидетельствует о том, что физиологические свойства микроорганизмов надо изучать не на изолированных искусственных средах, а в естественной среде их обитания. На процессы окисления аммиака влияют не только микробы, но и их ферменты. Кроме органического вещества, на нитрификацию оказывает влияние концентрация аммиака. Его действие на культуру резко проявляется в условиях жидких сред. В почве же аммиак, находится в адсорбированном состоянии и не может оказывать угнетающего действия. Поэтому нитробактер сразу же окисляет азотистую кислоту в азотную. На процесс нитрификации положительно сказывается присутствие кислорода. В обрабатываемых почвах процесс нитрификации протекает более интенсивно. Примерно также нитрификация проходит на черноземных почвах, особенно когда в них достаточное количество аммонифицирующих микробов, готовящих пищу (среду) для нитрификаторов. У солонцов меньшая нитрифицирующая способность. В почвах азотистая кислота не накапливается, поскольку Nitrosomonas и Nitrobacter встречаются в одной среде, находятся в своеобразном симбиозе. Нитрификаторы способны осуществлять хемосинтез, то есть создавать органическое вещество из углерода диоксида и воды за счет химической энергии окисления аммиака до азотистой кислоты и азотистой до азотной кислоты. Нитрификаторы чувствительны к кислой среде, они лучше развиваются при рН 8,3—9,3. В результате жизнедеятельности нитрифицирующих бактерий на 1 га почвы может накапливаться за год до 300 кг азотной кислоты. Д.Н. Прянишников установил, что одним из важнейших условий использования аммонийных и нитратных солей является реакция среды: в слабокислой среде при рН 5 лучше усваиваются нитраты, и, наоборот, в нейтральной среде при рН5лучше поглощаются аммонийные соли. Для использования аммонийных солей необходимо достаточное количество углеводов в растениях, без которых задерживается превращение их в амиды, накапливается аммиак, действующий токсический на растение. При питании растений физиологический нейтральной солью (NН4NO3) и реакции среды, близкой к нейтральной, лучше усваивается катион NН4+, чем анион NО3-, в этом случае соль будет физиологический кислой. Таким образом, реакция среды имеет большое значение для усвоения растениями соединений, содержащих азот. Экспериментально доказано, что преимущественное поглощение аммонийных солей свойственно растениям, склонным к усиленному образованию органических кислот.
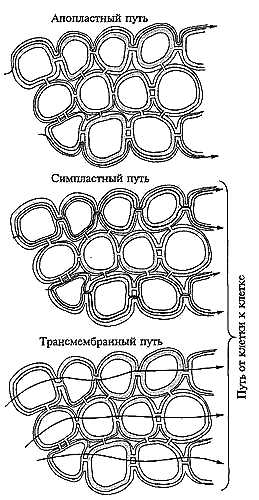
12. Химический состав растений, как известно, отражает элементный состав почв. Поэтому избыточное накопление ТМ растениями обусловлено, прежде всего, их высокими концентрациями в почвах. В своей жизнедеятельности растения контактируют только с доступными формами ТМ, количество которых, в свою очередь, тесно связано с буферностью почв. Однако способность почв связывать и инактивировать ТМ имеет свои пределы, и когда они уже не справляются с поступающим потоком металлов, важное значение приобретает наличие у самих растений физиолого-биохимических механизмов, препятствующих их поступлению. Механизмы устойчивости растений к избытку ТМ могут проявляться по разным направлениям: одни виды способны накапливать высокие концентрации ТМ, но проявлять к ним толерантность; другие стремятся снизить их поступления путем максимального использования своих барьерных функций. Для большинства растений первым барьерным уровнем являются корни, где задерживается наибольшее количества ТМ, следующий – стебли и листья, и наконец, последний ≈ органы и части растений, отвечающие за воспроизводительные функции. Уровень накопления ТМ разными растениями в зависимости от их генетических и видовых особенностей при одинаковом содержании ТМ в почвах. Отмечаются случаи, когда разные сорта одной культуры, произрастающие на одинаково загрязненной почве, содержали различные количество ТМ. Данный факт, по-видимому, обусловлен присущим всем живым организмам внутривидовым полиморфизмом, способным проявить себя и при техногенном загрязнении природной среды. Это свойство у растения может стать основой генетически – селекционных исследований с целью создания сортов с повышенными защитными возможностями по отношению к избыточным концентрациям ТМ. Несмотря на существенную изменчивость различных растений к накоплению ТМ, биоаккумуляция элементов имеет определенную тенденцию, позволяющую упорядочить их в несколько групп: 1) Cd, Cs, Rb – элементы интенсивного поглощения; 2) Zn, Mo, Cu, Pb, As – средней степени поглощения; 3) Mn, Ni – слабого поглощения; 4) Se, Fe, Ва – элементы, труднодоступные растениям. Другой путь поступления ТМ в растения ≈ некорневое поглощение из воздушных потоков. Оно имеет место при значительном выпадении металлов из атмосферы на листовой аппарат, чаще всего вблизи крупных промышленных предприятий. Поступление элементов в растения через листья (или фолиарное поглощение) происходит, главным образом, путем неметаболического проникновения через кутикулу. ТМ, поглощенные листьями, могут переноситься в другие органы и ткани и включаться в обмен веществ. Токсичность тяжелых металлов связана с их физико-химическими свойствами, со способностью к образованию прочных соединений с рядом функциональных группировок на поверхности и внутри клеток. Таким образом, действие металлов на растительный организм зависит от природы элемента, содержание его в окружающей среде, характера почвы, формы химического соединения, срока от момента загрязнения. Формирование химического состава растительного организма определяется биохимическими особенностями различных видов организмов, их возрастом и биохимическими закономерностями связи между элементами в организме. Содержание одних и тех химических элементов в различных частях растений. Важную роль в защите растений от избытка поступающих из почвы в корни ТМ выполняет корневая система. Задерживая избыточные ионы, корни тем самым способствуют сохранению в наземных органах невредных (благоприятных) концентрации химических элементов. Связывание ТМ органическими соединениями не всегда означает перевод в малоподвижное состояние. Отмечается, что некоторые комплексы ТМ с органическими лигандами способны проходить через мембраны легче, чем ионы этих металлов, и затем внутри клетки распадаться. При сильном загрязнении среды обитания поток ТМ становится столь большим, что может наблюдаться повышенное содержание их не только в вегетативных органах, но и в органах запаса ассимилятов. Растения выглядят угнетенными (хлорозы, некрозы), снижается их продуктивность. Это указывает на нарушение течения метаболических процессов. При очень сильном загрязнении среды обитания растения прекращают развитие, гибнут. Растения загрязняются также с поверхности в результате оседания из воздуха на листья и стебли металлосодержащих частиц. Поверхностное загрязнение может быть значительным. Накопление ТМ растениями, произрастающими на загрязненных почвах, в значительной степени зависит от уровня загрязнения. Однако сильная прямая корреляция между этими показателями обнаруживается не всегда, поскольку поток ТМ из почвы в растения определяется не только валовым содержанием, но и концентрацией в почве их подвижной формы, что тесно связано с химическим составом техногенных выбросов, защитными (буферными) возможностями почвы. Вблизи завода (0- 1,5 км) накопление ТМ столь велико, что возделывание сельскохозяйственных культур опасно для здоровья человека и животных. Свинец в растения в основном поступает через корни, тем не менее, он способен поглощаться и через листья. Накопление атмосферного свинца у растений вблизи автомобильных дорог может достигать 40% от его количества в растениях. Основное количество свинца сосредоточено в вегетативных органах, тогда как в репродуктивных органах накапливается 4-7% от его количества в растениях. Под влиянием марганца целый ряд почвенных микроорганизмов, участвующих в усвоении растениями атмосферного азота, усиливают свою активность. Среднее содержание марганца в растениях равно 0,00 1%. Марганец служит катализатором процессов дыхания растений, принимает участие в процессе фотосинтеза. Марганец входит в состав либо является активатором ряда ферментативных систем; регулирует отношение железа, тем самым влияя на окислительно-восстановительные процессы, совершающиеся с помощью железа. Марганец усиливает гидролитические процессы, в результате чего нарастает количество аминокислот, способствует продвижению ассимилятов, образующихся в процессе фотосинтеза от листьев к корням и другим органам. Токсичные для растений концентрации ТМ в зависимости от свойств почв могут варьировать в значительной степени. Эти уровни колеблются в 2-5 раз для различных ТМ при выращивании сельскохозяйственных культур. Критические уровни содержания ТМ в сухих растениях и растительных кормах (мг/кг вещества): для свинца -10-20 и 10-30; для никеля 26-154. Уровень содержания ТМ в почве, при котором происходит снижение продуктивности растений в 5-6 раз, называется токсичным. Наряду с физиологическими системами, ограничивающими поступление ТМ, растения располагает мощным аппаратом, выводящим их при выделении метаболитов. От избыточного количества ТМ растения могут освобождаться с корневыми выделениями, в процессе транспирации и дыхания. Растения транспортируют вместе с влагой значительное количество ТМ, составляющее целые проценты от их содержания в растительном организме. Проблема избыточного накопления ТМ в растениях затрагивает широкий круг вопросов, касающихся механизмов поглощения, транспорта, метаболизма и распределения его в органах и тканях. Биогенные элементы играют существенную роль в обеспечении физико-биохимических барьеров, при проникновении ТМ в растения. Избыточные концентрации ТМ отрицательно влияют на синтез и функции многих биохимических активных соединений: ферментов, витаминов, пигментов и др. Повышенные по сравнению с фоном концентрации ТМ существенно влияют на фотосинтетическую деятельность растений. При изучении белкового обмена растений показано, что избыточная концентрация ТМ влияет на уменьшение содержания альбуминов и возрастает количество проламинов, глютелинов и нерастворимого остатка. Это указывает на снижение содержания лизина. Растения обладают способностью ограничивать накопление элементов – загрязнителей в надземных органах, особенно в органах запасания ассимилятов. Растения являются хорошими индикаторами, позволяющими определить степень загрязнения почв соединениями тяжелых металлов. Степень накопления соединений тяжелых металлов в растениях определяется уровнем загрязнения почв, расстоянием от техногенного источника, количеством атмосферных выпадений и видовыми особенностями растений. При этом повышенные концентрации соединений ТМ содержатся в корнях и листьях. Содержание соединений ТМ в растениях сельскохозяйственных районов в семь раз меньше, чем в растениях, растущих в зоне городов. В связи с тем, что многие растения используются в пищу или корма возникает проблема взаимоотношения растений и тяжелых металлов в окружающей среде, она является актуальной и требует дальнейшего ее изучения. Оценка уровней безопасного загрязнения почв ТМ проводится исходя из недопустимости повышения порога адаптационной возможности наиболее чувствительной группы населения и экологической адаптационной способности почвы. Почва служит естественным барьером на пути ТМ и сдерживает их поступление в растения и сопредельные среды. Предельная допустимая концентрация в почвах для РЬ-фон +20 мг/кг. Сохранить почву в современных условиях практически невозможно, так как вся поверхность земного шара подвержена техногенному воздействию. Наиболее мощные потоки ТМ возникает вокруг предприятий черной и цветной металлургии. Исключение составляет свинец, в окружающую среду в большом количестве поступает также от автотранспорта. Урбанизация общества приводит к тому, что в почвах городов значительно повышается содержание. 13. Основным источником влаги является вода, находящаяся в почве, и основным органом поглощения воды — корневая система. Роль этого органа, прежде всего, заключается в том, что благодаря огромной поверхности обеспечивается поступление воды в растение из большего объема почвы. Сформировавшаяся корневая система представляет собой сложный орган с хорошо дифференцированной внешней и внутренней структурой. Определение размеров корневых систем требует специальных методов. Очень много в этом отношении достигнуто благодаря работам русских физиологов В. Г. Ротмистрова, А. П. Модестова, И. В. Красовской. Оказалось, что общая поверхность корней обычно превышает поверхность надземных органов в 140-150 раз. Подсчитано, что число корней у однолетних сеянцев яблони достигает 45 тыс. Корневые системы даже однолетних хлебных злаков проникают в почву на глубину 1,5-2 м. При выращивании одиночного растения ржи было установлено, что общая длина его корней может достигать 600км, при этом на них образуется 15 млрд. корневых волосков. Эти данные говорят об огромной потенциальной способности к росту корневых систем. Однако при росте растений в фитоценозах, с достаточно большой густотой их стояния, размеры корневых систем заметно уменьшаются. Рост корня, его ветвление продолжаются в течение всей жизни растительного организма, т. е. практически он не ограничен. Меристемы — образовательные ткани — расположены на верхушке каждого корня. Доля меристематических клеток сравнительно велика (10% по массе против 1% у стебля). Рост корней отличается большой скоростью. Считается, что одно растение риса в благоприятных условиях может образовать до 5 км новых корней в сутки. За счет этого прироста корневой системы в растение может дополнительно поступать 1,5 л воды. Только благодаря такому интенсивному росту корневые системы растения могут использовать скудно рассеянную в почве воду. Важное значение имеет явление гидротропизма, при котором рост корневой системы как бы идет из более иссушенных слоев почвы к более влажным. В зависимости от типа растений распределение корневой системы в почве различно. У некоторых растений корневая система проникает на большую глубину, у других главным образом распространяется в ширину. С физиологической точки зрения корневая система неоднородна. Далеко не вся поверхность корня участвует в поглощении воды. В каждом корне различают несколько зон, правда, не всегда все зоны выражены одинаково четко. Окончание корня снаружи защищено корневым чехликом, напоминающим округлый колпачок, состоящий из живых тонкостенных продолговатых клеток. Корневой чехлик служит защитой для точки роста. Клетки корневого чехлика слущиваются, что уменьшает трение и способствует проникновению корня в глубь почвы. Под корневым чехликом расположена меристематическая зона. Меристема состоит из многочисленных мелких, усиленно делящихся, плотно упакованных клеток, почти целиком заполненных протоплазмой. Следующая зона — зона растяжения. Здесь клетки увеличиваются в объеме (растягиваются). Одновременно в этой зоне появляются дифференцированные ситовидные трубки. Затем следует зона корневых волосков. При дальнейшем увеличении возраста клеток, а также расстояния от кончика корня корневые волоски исчезают, начинается кутинизация и опробковение клеточных оболочек. Поглощение воды происходит главным образом клетками зоны растяжения и зоны корневых волосков. Некоторое количество воды может поступать и через опробковевшую зону корня. Это главным образом наблюдается у деревьев. В этом случае вода проникает через чечевички или поранения. Поверхность корня в зоне корневых волосков покрыта ризодермой. Это однослойная ткань с двумя видами клеток, формирующими и не формирующими корневые волоски. В настоящее время показано, что клетки, формирующие корневые волоски, отличаются особым типом обмена веществ. Корневые волоски растут путем растяжения клеточной оболочки, которое происходит с большой скоростью (0,1 мм/ч). Для их роста очень важно присутствие кальция. У большинства растений клетки ризодермы обладают тонкими стенками. Вслед за ризодермой до перицикла идут клетки коры. Кора состоит из нескольких слоев паренхимных клеток. Важной особенностью коры является развитие системы крупных межклетников. На границе коры и центрального цилиндра развивается один слой плотно прилегающих друг к другу клеток — эндодерма, для которой характерно наличие поясков Каспари. Цитоплазма в клетках эндодермы плотно прилегает к клеточным оболочкам. По мере старения вся внутренняя поверхность клеток эндодермы, за исключением пропускных клеток, покрывается суберином. При дальнейшем старении сверху могут накладываться еще слои. По-видимому, именно клетки эндодермы служат основным физиологическим барьером для передвижения как воды, так и питательных веществ. В центральном цилиндре расположены проводящие ткани корня. При рассмотрении структуры корня в продольном направлении важно отметить, что начало роста корневых волосков, появление поясков Каспари в стенках эндодермы и дифференциация сосудов ксилемы происходят на одном и том же расстоянии от апикальной меристемы. Именно эта зона является зоной снабжения растений питательными веществами. Обычно поглощающая зона составляет около 5 см в длину. Величина ее зависит от скорости роста корня в целом. Чем медленнее растет корень, тем зона поглощения короче. Движение воды в радиальном направлении из почвенного раствора в проводящие элементы ксилемы осуществляется как непосредственно через клетки, так и по апопласту тканей корня (рис. 1). Напомним, что под термином Апопласт У растений понимают единую систему клеточных стенок и межклеточных полостей. Транспорт воды через клетки Получил название "От клетки к клетке" («Cell To Cell»). В этом случае, преодолевая последовательно расположенные в радиальном направлении ткани корня, вода при движении от клетки к клетке может пересекать мембраны или идти по непрерывному цитоплазматическому пути через соединяющие соседние клетки плазмодесмы. Единая система, объединяющая с помощью плазмодесм протопласты растительных клеток, называется Симпластом. В соответствии с этим путь движения воды от клетки к клетке подразделяется на трансмембранный и симпластный. При трансмембранном движении вода в пределах одной клетки дважды пересекает плазмалемму и клеточную стенку, а если заходит в вакуоль, то дважды пересекает и тонопласт. В трансмембранном движении воды важную роль играют образованные аквапоринами водные каналы, которые характеризуются высокой водопроницаемостью. Таким образом, движение воды от клеток эпидермиса к сосудам ксилемы может осуществляться по трем параллельным направлениям: 1) через клеточные мембраны (трансмембранный путь); Эти три пути движения воды не автономны: вода может перемещаться по комбинированным траекториям, например пройти некоторый путь по апопласту, затем пересечь ПМ и далее передвигаться от клетки к клетке по симпласту.
Надо отметить, что в целом корневые системы значительно менее разнообразны по сравнению с надземными органами, в связи с тем, что среда их обитания более однородна. Это не исключает того, что корневые системы изменяются под влиянием тех или иных условий. Хорошо показано влияние температуры на формирование корневых систем. Как правило, оптимальная температура для роста корневых систем несколько ниже по сравнению с ростом надземных органов того же растения. Все же сильное понижение температуры заметно тормозит рост корней и способствует образованию толстых, мясистых, мало ветвящихся корневых систем. Большое значение для формирования корневых систем имеет влажность почвы. Распределение корней по горизонтам почвы часто определяется распределением воды в почве. Обычно в первый период жизни растительного организма корневая система растет чрезвычайно интенсивно и, как следствие, скорее достигает более влажных слоев почвы. Некоторые растения развивают поверхностную корневую систему. Располагаясь близко к поверхности, сильно ветвящиеся корни перехватывают атмосферные осадки. В засушливых районах часто глубоко и мелко укореняющиеся виды растений растут рядом. Первые обеспечивают себя влагой за счет глубоких слоев почвы, вторые за счет усвоения выпадающих осадков. Важное значение для развития корневых систем имеет аэрация. Именно недостаток кислорода является причиной плохого развития корневых систем на заболоченных почвах. Растения, приспособленные к росту на плохо аэрируемых почвах, имеют в корнях систему межклетников, которые вместе с межклетниками в стеблях и листьях составляют единую вентиляционную систему. Большое значение имеют условия питания. Показано, что внесение фосфорных удобрений способствует углублению корневых систем, а внесение азотных удобрений — их усиленному ветвлению. 14. Степень глубины физиологического покоя и его длительность неодинаковы. Семена выводятся из состояния покоя различным образом. Некоторые семена, особенно однолетних растений, легко набухают и прорастают уже под влиянием увлажнения. Для прорастания других и нормального развития проростка обязательна холодная стратификация, т.е. длительное выдерживание их при пониженной температуре, во влажной среде и в условиях хорошей аэрации. Наконец, существует еще одна группа так называемых "твердосеменных", семенная кожура которых в силу ее структурных особенностей водонепроницаема. Такие семена прорастают только после скарификации - искусственного нарушения целостности кожуры с помощью надцарапывания, перетирания с песком, ошпаривания кипятком и т.д. В природе такие семена набухают и прорастают обычно под влиянием резкой смены температурных режимов, способствующих нарушению целостности оболочки. Прорастанием семян называют их переход от состояния покоя к вегетативному росту зародыша и формированию из него проростка. Прорастание начинается при оптимальном для каждого вида сочетании влажности и температуры среды, при свободном доступе кислорода. Прорастание семян сопровождается сложными биохимическими и морфофизиологическими процессами. При поступлении воды в семенах резко усиливается процесс дыхания, активизируются ферменты, запасные вещества переходят в легкоусвояемую, подвижную форму, образуются полирибосомы и начинается синтез белка и других веществ. Рост зародыша обычно начинается с прорыва покровов удлиняющимися зародышевым корнем игипокотилем в области микропилярного следа. После появления корня почечка развивается в побег, на котором развертываются настоящие листья (рис. 51). В практике сельского хозяйства жизнеспособность и качество семян характеризуются всхожестью, т.е. процентом семян, давших нормальные проростки в оптимальных для них условиях за определенный срок. Для полевых культур этот срок равен 6-10 суткам, для древесных - до 2 месяцев. 15. Солеустойчивость растений — это способность растений противостоять засолению, не снижая интенсивность течения основных физиологических процессов. Изучение солеустойчивости растений имеет большое практическое значение, поскольку океаны, воды которых содержат 3 — 4% солей, занимают около 75% поверхности Земли, более четверти всех почв засолены, а еще одна треть всех почв имеет тенденцию к засолению. Согласно Б. П. Строганову (1962), по степени засоления различают незасоленные, слабозасоленные, средне-засоленные почвы и солончаки. Тип засоления определяется по содержанию анионов в почве: хлоридное, сульфатное, сульфатно-хлоридное, хлоридно-сульфатное и карбонатное. Преобладающим катионом в таких почвах является натрий, но встречается также карбонатно-магниевое, хлоридно-магниевое засоление, Растения, приспособленные к существованию в условиях избыточного засоления, называют галофитами. В чем же вред засоления? Засоление приводит к созданию в почве низкого (отрицательного) водного потенциала, поэтому поступление воды в растение сильно затруднено. Под влиянием солей происходит нарушение ультраструктуры клеток, в частности изменение в структуре хлоропластов. Особенно это проявляется при хлоридном засолении. Вредное влияние высокой концентрации солей связано с повреждением мембранных структур, в частности плазмалеммы, вследствие чего возрастает ее проницаемость, теряется способность к избирательному накоплению веществ. В этом случае соли поступают в клетку пассивно, и это усиливает повреждение клетки. На засоленных почвах большая концентрация натрия препятствует накоплению других катионов, в том числе кальция. Высокая концентрация солей нарушает азотный обмен (накапливается аммиак), возникают признаки серного голодания. Наоборот, в условиях засоления, связанного с высокой концентрацией серно-кислых солей, наблюдается обратный процесс — избыточное накопление серы, что также приводит к синтезу и накоплению ядовитых соединений. Повышенная концентрация хлоридных солей может действовать как разобщитель процессов окисления и фосфорилирования, т. е. снижается процесс синтеза АТФ в дыхании. Следует отметить, что отрицательное действие высокой концентрации солей сказывается прежде всего на функционировании корневой системы. При этом в корнях страдают наружные клетки, непосредственно соприкасающиеся с раствором соли. В стебле наиболее подвержены действию солей клетки проводящей системы. В основе устойчивости растений к солям лежат различные физиологические механизмы. К первой группе относятся механизмы, запускающие реакции обмена веществ, которые нейтрализуют неблагоприятное действие солей. Примером может служить окислительное разрушение токсичных соединений серы и ее производных, а также накопление веществ, регулируемых осмотические свойства клеток и оказывающих защитное влияние, например, аминокислота пролин. Важное значение в процессе приспособления к засолению имеет накопление веществ, относящихся к полиаминам (путресцин, спермидин и др.). В определенных концентрациях эти вещества оказывают защитное действие. Это связано, по-видимому, с тем, что они стабилизируют структуру нуклеиновых кислот и повышают устойчивость растений. Ко второй группе относятся механизмы, регулирующие транспорт ионов из среды в клетку. Этот тип приспособлений связан с защитными функциями мембран. Рассмотрим отдельные группы галофитов и их приспособления противостоять высоким концентрациям солей. Все гало-фиты можно разделить на три группы: 1. Настоящие галофиты (эвгалофиты) — наиболее солеустойчивые растения, накапливающие в ва
|
||
|
|
Последнее изменение этой страницы: 2016-08-01; просмотров: 957; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.214 (0.018 с.) |