Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
III. центральная нервная системаСодержание книги
Поиск на нашем сайте
Филогенез центральной нервной системы Нервные клетки кишечнополостных соединены между собой в единую сеть («диффузная» нервная система). В связи с этим, в ответную реакцию на раздражение вовлекается весь организм. В процессе последующей эволюции происходит концентрация нервных клеток и образование нервных центров. У плоских червей, в связи с дифференцировкой переднего и заднего конца тела, происходит концентрация нервных клеток и образование нервного центра в виде головного ганглия или окологлоточного нервного кольца. От них отходят вдоль тела нервные стволы, состоящие из нервных волокон и клеток. У кольчатых червей нервная система состоит из парного головного ганглия и брюшной нервной цепочки, представленной двумя нервными стволами, идущими по брюшной стороне и образующими в каждом сегменте по нервному узлу. Каждая пара узлов соединена между собой поперечными связями (лестничный тип нервной системы). В переднем отделе слияние ганглиев приводит к значительному укрупнению и усложнению строения головного ганглия (головного мозга). У хордовых возникает принципиально новый тип нервной системы в виде нервной трубки. У позвоночных ее передний конец прогрессивно развивается и превращается в головной мозг. Гомология нервной системы позвоночных и нервной системы низших хордовых проявляется уже при закладке отделов головного мозга в эмбриональном периоде. У всех классов позвоночных закладка нервной системы последовательно проходит стадии нервной пластинки, желоба и нервной трубки с полостью невроцелем. Затем начинается период дифференцировки нервной трубки на головной и спинной мозг, когда прямая до сих пор трубка образует ряд вздутий, изгибов и дает утолщения стенки в определенных местах. Формирование головного мозга у всех позвоночных начинается с образования на переднем конце трубки трех вздутий, или мозговых пузырей. Первый из них носит название переднего (prosencephalon), второй – среднего (mesencephalon) и третий – ромбовидного (заднего) мозгового пузыря (rhombencephalon). После стадии трех мозговых пузырей следует стадия пяти мозговых пузырей. Они дают начало дефинитивным отделам мозга. Передний мозговой пузырь делится поперечной перетяжкой на два отдела. Первый их них (передний) образует передний отдел головного мозга (telencephalon), который у большинства позвоночных образует так называемые полушария большого мозга. Из задней части переднего мозгового пузыря развивается промежуточный мозг (diencephalon). Средний мозговой пузырь не делится и целиком преобразуется в средний мозг (mesencephalon). Задний мозговой пузырь также подразделяется на два отдела: в передней части он образует задний мозг (metencephalon), а из заднего отдела формируется продолговатый мозг (meduiia oblongata), который без резкой границы переходит в спинной мозг.
Таким образом, у всех классов позвоночных головной мозг состоит из пяти основных отделов: переднего, промежуточного, среднего, заднего и продолговатого, расположенных в одной и той же последовательности. Степень развития отделов у различных классов не одинакова. В процессе образования пяти пузырей невроцель образует ряд расширений, которые носят названия желудочков мозга. Желудочки соответствуют отделам мозга и сообщаются между собой и со спинномозговым каналом. В каждом отделе мозга различают крышу, или мантию (pallium), и дно, или основание. Крышу составляют части мозга, лежащие над желудочками, дно (основание) лежит под желудочками. Вещество мозга неоднородно. Скопления нервных клеток имеют более темную окраску – серое вещество, скопление отростков нервных клеток более светлую – белое вещество. Слой серого вещества (нервных клеток) на поверхности крыши любого отдела мозга носит название коры (cortex). Скопления серого вещества в толще белого называются ядрами.
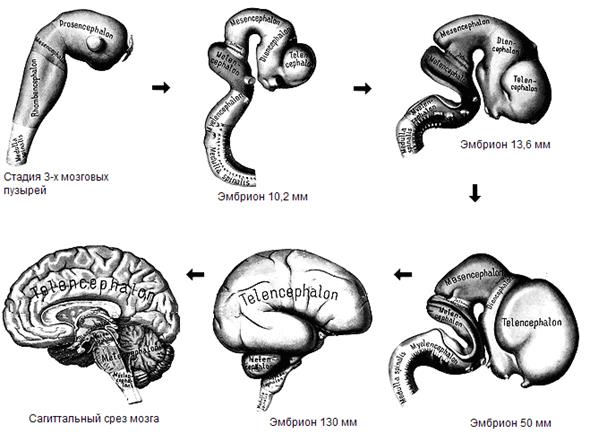
Онтогенез центральной нервной системы Эмбриогенез головного мозга начинается с развития в передней (ростральной) части мозговой трубки двух первичных мозговых пузырей, возникающих в результате неравномерного роста стенок нервной трубки (архэнцефалон и дейтерэнцефалон). Дейтерэнцефалон, как и задняя часть мозговой трубки (впоследствии спинной мозг), располагается над хордой. Архэнцефалон закладывается впереди нее. Затем в начале четвертой недели у зародыша дейтерэнцефалон делится на средний (mesencephalon) и ромбовидный (rhombencephalon) пузыри. А архэнцефалон превращается на этой (трехпузырной) стадии в передний мозговой пузырь (prosencephalon). В нижней части переднего мозга выпячиваются обонятельные лопасти (из них развиваются обонятельный эпителий носовой полости, обонятельные луковицы и тракты). Из дорсолатеральных стенок переднего мозгового пузыря выступают два глазных пузыря. В дальнейшем из них развиваются сетчатка глаз, зрительные нервы и тракты.
Рис. 1. Онтогенетические этапы развития головного мозга
На шестой неделе эмбрионального развития передний и ромбовидный пузыри делятся каждый на два, и наступает пятипузырная стадия. Передний пузырь – конечный мозг – разделяется продольной щелью на два полушария. Полость также делится, образуя боковые желудочки. Мозговое вещество увеличивается неравномерно, и на поверхности полушарий образуются многочисленные складки – извилины, отделенные друг от друга более или менее глубокими бороздами и щелями. Каждое полушарие разделяется на четыре доли, в соответствие с этим полости боковых желудочков делятся также на 4 части: центральный отдел и три рога желудочка. Из мезенхимы, окружающей мозг зародыша, развиваются оболочки мозга. Серое вещество располагается и на периферии, образуя кору больших полушарий, и в основании полушарий, образуя подкорковые ядра. Задняя часть переднего пузыря остается неразделенной и называется теперь промежуточным мозгом. Функционально и морфологически он связан с органом зрения. На стадии, когда границы с конечным мозгом слабо выражены, из базальной части боковых стенок образуются парные выросты – глазные пузыри, которые соединяются с местом их происхождения при помощи глазных стебельков, впоследствии превращающихся в зрительные нервы. Наибольшей толщины достигают боковые стенки промежуточного мозга, которые преобразуются в зрительные бугры, или таламус. В соответствии с этим полость III желудочка превращается в узкую сагиттальную щель. В вентральной области (гипоталамус) образуется непарное выпячивание – воронка, из нижнего конца которой происходит задняя мозговая доля гипофиза – нейрогипофиз. Третий мозговой пузырь превращается в средний мозг, который развивается наиболее просто и отстает в росте. Стенки его утолщаются равномерно, а полость превращается в узкий канал – Сильвиев водопровод, соединяющий III и IV желудочки. Из дорсальной стенки развивается четверохолмие, а из вентральной – ножки мозга. Ромбовидный мозг делится на задний и добавочный. Из заднего формируется мозжечок – сначала червь мозжечка, а затем полушария, а также мост. Добавочный мозг превращается в продолговатый мозг. Стенки ромбовидного мозга утолщаются – как с боков, так и на дне, только крыша остается в виде тончайшей пластинки. Полость превращается в IV желудочек, который сообщается с Сильвиевым водопроводом и с центральным каналом спинного мозга. В результате неравномерного развития мозговых пузырей мозговая трубка начинает изгибаться (на уровне среднего мозга – теменной прогиб, в области заднего мозга – мостовой и в месте перехода добавочного мозга в спинной – затылочный прогиб). Теменной и затылочный прогибы обращены наружу, а мостовой – внутрь. Структуры головного мозга, формирующиеся из дейтерэнцефалона: средний, задний и добавочный мозг – составляют основу ствола головного мозга (truncus cerebri). Он является ростральным продолжением спинного мозга и имеет с ним общие черты строения. Проходящая по латеральным стенкам спинного мозга и стволового отдела головного мозга парная пограничная борозда (sulcus limitons) делит мозговую трубку на основную (вентральную) и крыловидную (дорзальную) пластинки. Из основной пластинки формируются моторные структуры (передние рога спинного мозга, двигательные ядра черепно-мозговых нервов). Над пограничной бороздой из крыловидной пластинки развиваются сенсорные структуры (задние рога спинного мозга, сенсорные ядра ствола мозга), в пределах самой пограничной борозды – центры вегетативной нервной системы.
Спинной мозг Спинной мозг, medulla spinalis, располагается в позвоночном канале. Он начинается на уровне края большого затылочного отверстия и заканчивается на уровне верхнего края II поясничного позвонка. Внешнее строение. Особенности внешнего строения характеризуются наличием следующих структурных образований: • Шейное утолщение, intumescentia cervicalis, расположенное в проекции III шейного – II грудного позвонков. • Пояснично-крестцовое утолщение, intumescentia lumbosacralis, расположенное в проекции IX грудного – II поясничного позвонков. • Мозговой конус, conus medullaris, – каудальный конец спинного мозга, расположенный в проекции I-II поясничных позвонков. • Концевая (терминальная) нить, filum terminale, заканчивается в надкостнице копчиковых позвонков. • Передняя срединная щель, fissura mediana anterior, – глубокая продольная щель на передней поверхности спинного мозга. • Задняя срединная борозда, sulcus medianus posterior, – менее выраженная продольная борозда на его задней поверхности. (Передняя срединная щель и задняя срединная борозда разделяют спинной мозг на две симметричные половины.) • Задняя боковая борозда, sulcus posteriolateralis, проходит параллельно задней срединной борозде. • Задние корешковые нити, fila radicularia posteriores, вступают в заднюю боковую борозду (в совокупности – задний корешок) – чувствительные; • Передняя боковая борозда, sulcus anteriolateralis, проходит параллельно передней срединной щели. • Передние корешковые нити, fila radicularia anteriores, выходят из передней боковой борозды (в совокупности – передний корешок) – двигательные. • Спинномозговой узел, ganglion spinale, – небольшое утолщение по ходу заднего корешка, образованное телами чувствительных псевдоуниполярных нейронов. • Спинномозговой нерв, nervus spinalis, образуется при слиянии переднего и заднего корешков. Каждый спинномозговой нерв покидает позвоночный канал через соответствующее межпозвоночное отверстие. Спинной мозг состоит из 31 cпинномозгового сегмента.
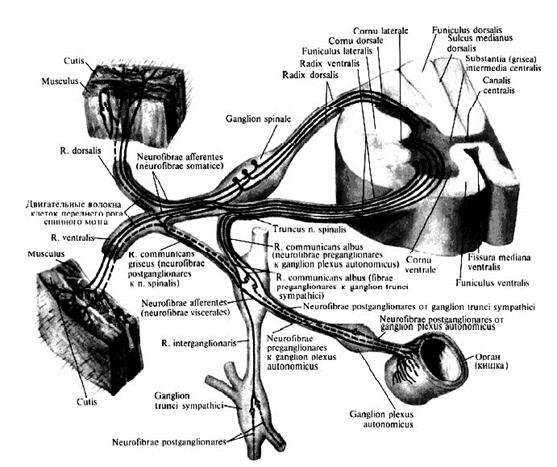
Спинномозговой (спинальный) сегмент – участок спинного мозга, корешки которого формируют одну пару спинномозговых нервов (рис. 2).
Рис. 2. Ход волокон спинномозговых нервов и их связь с симпатическим стволом. Различают следующие сегменты: 8 шейных, 12 грудных, 5 поясничных, 5 крестцовых, 1 копчиковый. Так как длина спинного мозга меньше длины позвоночного канала, то его сегменты лежат несколько выше соответствующих им позвонков. Это несоответствие между сегментами и позвонками различно на разных уровнях и наиболее сильно выражено в нижних отделах спинного мозга.
Ниже второго поясничного позвонка в позвоночном канале находятся только корешки поясничных и крестцовых нервов. Они проходят почти параллельно продольной оси спинного мозга, так что мозговой конус и терминальная нить оказываются расположенными внутри густого пучка нервных корешков, который получил название конского хвоста, cauda equina. Внутреннее строение. На поперечном разрезе спинного мозга серое вещество, которое окружает центральный канал, представлено парными передними, задними, и боковыми рогами (С8-L2), которые в действительности представляют собой непрерывные столбы (columnae griseae), идущие вдоль спинного мозга. Рога серого вещества обеих половин спинного мозга соединяются с помощью передней серой спайки и задней серой спайки, спереди от которой располагается передняя белая спайка. В сером веществе спинного мозга тела нервных клеток, сходных по строению и функции, объединяются, образуя ядра. В переднем роге находится группа так называемых двигательных ядер, которые состоят преимущественно из мотонейронов (крупных мультиполярных нейронов). Аксоны мотонейронов покидают спинной мозг в составе передних корешков, затем в составе смешанных спинномозговых нервов направляются к соматическим мышцам. Описано 6 ядер переднего рога, которые соответственно положению получили названия: переднемедиальное, заднемедиальное, переднелатеральное, заднелатеральное, центральное и зацентральное. Ядра заднего рога образованы главным образом вставочными нейронами и включают: • студенистое вещество, substantia gelatinosa, – образовано телами ассоциативных нейронов переднего спинноталамического пути; • собственные ядра, nuclei proprii, – образованы телами ассоциативных нейронов латерального спинно-таламического пути; • грудное ядро, nucleus thoracicus (столб Кларка–Штиллинга, columna Stilling-Clarke) (C8-L2), – образовано телами ассоциативных нейронов заднего спинномозжечкового пути; • губчатая зона, zona spongiosa, – получила свое название в связи с наличием в этом отделе крупнопетлистой глиальной сети, содержащей нервные клетки; • центральное промежуточное вещество, substantia intermedia centralis, – отростки клеток этого вещества участвуют в образовании спинно-мозжечкового пути. В промежуточной зоне серого вещества находятся: • латеральное промежуточное ядро, nucleus intermediolatemlis (C8-L2), – центр симпатической нервной системы;
• медиальное промежуточное ядро, nucleus intermediomedialis, – образовано телами ассоциативных нейронов переднего спинно-мозжечкового пути. Белое вещество спинного мозга окружает серое вещество и разделяется на симметричные задние, боковые и передние канатики, funiculus anterior, lateralis et posterior. Оно образовано миелиновыми нервными волокнами, которые группируются в проводящие пути. Различают нисходящие (двигательные, эфферентные) и восходящие (чувствительные, афферентные) пути. В задних канатиках располагается афферентный путь Голля и Бурдаха, в передних канатиках проходят эфферентные передний пирамидный, покрышечно-спинномозговой и преддверно-спинномозговой пути, а в боковых канатиках – как афферентные, так и эфферентные проводящие пути. Оболочки спинногомозга. Спинной мозг окружен твердой, dura mater spinalis, паутинной, arachnoidea spinalis, и мягкой, pia mater spinalis, оболочками. Твердая мозговая оболочка образована плотной волокнистой соединительной тканью и образует просторный футляр, который простирается от уровня большого затылочного отверстия до II крестцового позвонка. Между твердой оболочкой спинного мозга и надкостницей позвонков находится эпидуральное пространство, cavitas epiduralis, заполненное рыхлой волокнистой соединительной тканью и жировой клетчаткой, содержащей большое число лимфатических сосудов и густое венозное сплетение. Внутренняя поверхность твердой мозговой оболочки отделяется от паутинной оболочки капиллярным субдуральным пространством, spatium subdurale. Тонкая бессосудистая паутинная оболочка лежит между твердой и мягкой оболочками и отделена от последней субарахноидальным пространством, cavitas subarachnoidalis, в котором циркулирует спинномозговая жидкость. Эта полость с помощью зубчатой связки делится на переднюю и заднюю части. Мягкая мозговая оболочка плотно прилегает к спинному мозгу и сращена с ним. Она состоит из двух листков, между которыми находится так называемое интерпиальное пространство, где располагается сеть кровеносных сосудов.
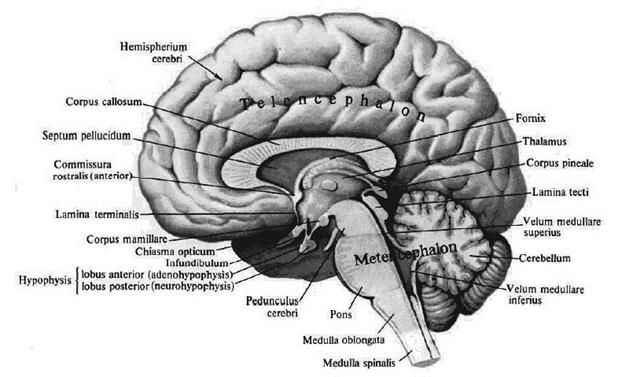
Головной мозг Головной мозг, с окружающими его оболочками находится в полости мозгового черепа. Верхнелатеральная поверхность головного мозга по форме соответствует внутренней вогнутой поверхности свода черепа. Нижняя поверхность – основание головного мозга, имеет сложный рельеф, соответствующий черепным ямкам внутреннего основания черепа. Масса мозга взрослого человека колеблется от 1100 до 2000 г. На протяжении от 20 до 60 лет масса и объем остаются максимальным и постоянным для каждого индивидуума. Основные части головного мозга: большой мозг, ствол и малый мозг (мозжечок). Полушария большого мозга разделены глубокой вертикальной щелью, fissura longitudinalis cerebri. Поперечная щель, fissura transversa cerebri, отделяет большой мозг от нижерасположенного мозжечка, cerebellum. На поверхности полушарий определяются борозды, sulci cerebri, между которыми расположены извилины, gyri cerebri.
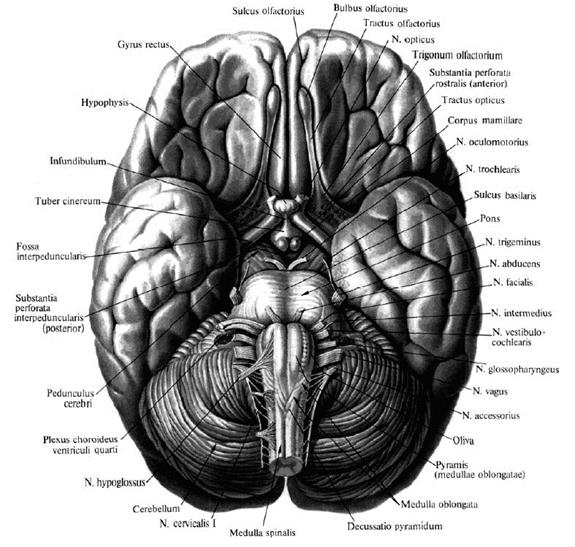
Рис. 3. Медиальная поверхность головного мозга. Вентральная (нижняя) поверхность называется основанием мозга. По направлению от лобного полюса к затылочному здесь можно увидеть следующие образования: • Обонятельная луковица, bulbus olfactorius. • Обонятельный тракт, tractus olfactorius. • Обонятельный треугольник, trigonum olfactorium, который ограничен наружной и внутренней обонятельными полосками, striae olfactoriae medialis et lateralis. • Перекрест зрительных нервов, chiasma opticum. Спереди от перекреста находятся зрительные нервы, n. opticus, сзади – зрительные тракты, tractus opticus. • Переднее продырявленное вещество, subsfantia perforata anterior, расположенное по бокам от перекреста и зрительных трактов. • Серый бугор, tuber cinereum, который вытягивается в отросток, напоминающий воронку, infundibulum; с его помощью основание мозга соединяется с гипофизом, hypophysis. • Ножки мозга, crus cerebri, находятся справа и слева от серого бугра. • Сосцевидные тела, corpora mamillaria, лежат позади серого бугра. • Межножковая ямка, fossa interpeduncularis, занимает пространство между ножками мозга и кзади от сосцевидных тел. • Заднее продырявленное вещество, substantia perforata posterior, образует дно межножковой ямки. • Мост, pons (Varolii), с расположенной посередине срединной бороздой, sulcus basilaris. Кнаружи мост суживается и затем, направляясь в стороны и назад, погружается в мозжечок, формируя его средние ножки, pedunculus cerebellaris medius. • Продолговатый мозг, medulla oblongata, лежит позади моста. По срединной линии продолговатого мозга проходит передняя срединная шель, ограниченная с каждой стороны белым валиком, пирамидой, pyramis medullae oblongatae. Кнаружи от пирамиды лежит продолговатое возвышение, олива, oliva.
Рис. 4. Головной мозг, вид снизу.
На основание мозга выходят корешки 12 пар черепных нервов: • Обонятельный нерв, пn olfactorii (I). Первую пару черепных нервов составляют центральные отростки обонятельных клеток, расположенных в слизистой оболочке носовой полости, – это так называемые обонятельные нити, fila olfactoria. Они проникают в полость черепа через продырявленную пластинку решетчатой кости и подходят к обонятельной луковице. • Зрительный нерв, п. opticus (II), образует с одноименным нервом противоположной стороны зрительный перекрест, chiasma opticum, и далее продолжается под названием tractus opticus. • Глазодвигательный нерв, п. oculomotorius (III),выходит на внутренней поверхности ножек мозга из одноименной борозды. • Блоковый нерв, n. trochlearis (IV), выходит из верхнего мозгового паруса и появляется на основании снаружи от ножки мозга. • Тройничный нерв, п. trigeminus (V), лежит у переднего края варолиева моста, на границе со средними ножками мозжечка (linea trigeminofacialis). Появляется двумя корешками: тонким – двигательным и толстым – чувствительным. • Отводящий нерв, п. abducens (VI), выходит на границе заднего края моста и пирамиды продолговатого мозга. • Лицевой нерв, п. facialis (VII), находится между задним краем моста и оливой, в так называемом мосто-мозжечковом углу (linea trigeminofacialis). • Преддверно-улитковый нерв, п. vestibulocochlearis (VIII), лежит в мосто-мозжечковом углу снаружи от лицевого нерва. • Языкоглоточный нерв, п. glossopharyngeus (IX), 5-6 корешков этого нерва находятся позади олив. • Блуждающий нерв, п. vagus (X), 10-12 корешков этого нерва лежат позади олив ниже предыдущей пары. • Добавочный нерв, п. accessorius (XI), выходит многими корешками на боковой поверхности продолговатого мозга и шейной части спинного мозга. • Подъязычный нерв, п. hypoglossus (XII), появляется на основании мозга между пирамидой и оливой.
ПРОДОЛГОВАТЫЙ МОЗГ Внешнее строение. В продолговатом мозге выделяют вентральную, дорсальную и боковые поверхности. На вентральной поверхности верхней границей продолговатого мозга является задний край варолиева моста. За нижнюю границу принимают место выхода корешков первой пары шейных спинномозговых нервов. Рельеф вентральной поверхности определяют следующие образования: • Передняя срединная щель. Пирамида, которая определяется с каждой стороны от передней срединной щели и образована волокнами корково-спинномозгового пути. Большая часть волокон этого пути переходит срединную линию (decussatio pyramidorum) и далее проходит в боковом канатике противоположной стороны спинного мозга. • Оливанаходится снаружи от пирамиды. • Корешки подъязычного нерва, расположенные между пирамидой и оливой. • Передняя боковая борозда, где локализуются корешки IX, X и XI пар черепных нервов. Верхней границей продолговатого мозга на дорсальной поверхности являются медуллярные полоски IV-го желудочка. Рельеф дорсальной поверхности определяют следующие образования: • Задняя срединная борозда, которая снизу замыкается тонким мозговым листком – задвижкой, obex. Под задвижкой идущий в глубине центральный канал переходит в четвертый желудочек. • Задняя промежуточная борозда,которая разделяет восходящие из спинного мозга нежный и клиновидный пучки. • Бугорки тонкого и клиновидного ядер, которыми заканчиваются одноименные пучки. • Задняя боковая борозда, ограничивает снаружи клиновидный пучок. Между передней и задней боковыми бороздами располагается боковая часть продолговатого мозга, которая продолжается в нижние ножки мозжечка. Внутреннее строение. Серое вещество продолговатого мозга представлено следующими образованиями: •Ядра черепных нервов: - Двигательные ядра подъязычного (XII) и добавочного (XI) нервов. - Двигательные и чувствительные соматические, а также парасимпатические ядра блуждающего (X) и языкоглоточного (IX) нервов. - Чувствительное ядро тройничного нерва, которое располагается на всем протяжении продолговатого мозга, опускаясь в верхние шейные сегменты спинного мозга. • Ядерный комплекс оливы. Его ядра относятся к экстрапирамидной системе и имеют связи с мозжечком и спинным мозгом. • Ядра тонкого (Голля) и клиновидного (Бурдаха) пучков, nucleus gracilis и nucleus cuneatus, расположенные в одноименных бугорках дорсальной поверхности продолговатого мозга, образованы телами ассоциативных нейронов восходящих проводящих путей проприоцептивной чувствительности коркового направления (пути Голля и Бурдаха). Аксоны вторых нейронов, выходящие из указанных ядер, переходят на противоположную сторону продолговатого мозга, образуя так называемую медиальную петлю мозгового ствола, lemniscus medialis. Перекрест нервных волокон в дорсальной части продолговатого мозга получает название перекреста медиальных петель, или чувствительного перекреста, decussatio lemniscorum. • Ретикулярная формация, formatio reticularis, образована переплетением волокон, идущих в различных направлениях, и расположенными между ними нервными клетками, которые образуют отдельные клеточные группы (ядра ретикулярной формации). Ретикулярная формация лежит в дорсальной части продолговатого мозга, моста, ножек мозга и простирается в каудальную часть промежуточного мозга. Ее рассматривают как ростральное продолжение межнейронных сетей спинного мозга, которые становятся значительно более интенсивными. Аксоны нервных клеток ретикулярной формации опускаются в спинной мозг, восходят к таламическому и гипоталамическому отделам промежуточного мозга, к коре полушарий. Ретикулярная формация представляет собой полифункциональную структуру. Она является интегративным центром ствола, во многом определяющим направление и интенсивность потоков сенсорной информации к высшим отделам мозга, а также степень нисходящих влияний на активность спинальных и стволовых нейронов. Ретикулярная формация участвует в регуляции рефлекторной двигательной активности. Ядра ретикулярной формации образуют основные «центры жизнеобеспечения» ствола, такие как дыхательный и сосудодвигательный. Дыхательный центр продолговатого мозга окружает nucl. solitarius и nucl. ambiquus. Он состоит из центра вдоха и центра выдоха. В ретикулярной формации моста располагаются «пневмотаксический» центр (центр регуляции внешнего дыхания (вдоха и выдоха)) и центр «апноэ» (задержки дыхания). «Прессорный» и «депрессорный» сосудодвигательные центры обеспечивают рефлекторную регуляцию кровяного давления, «акселераторный» и «ингибиторный» центры, регулирующие частоту и силу сердечных сокращений. Ретикулярная формация участвует в модуляции болевой чувствительности. Установлено, что стимуляция определенных областей сетчатой субстанции может заметно уменьшить или прервать афферентную импульсацию. Ретикулярная формация обладает выраженным активизирующим действием на активность нейронов коры больших полушарий. Восходящие пути из ретикулярной формации охватывают широкие корковые поля. Они получили название «второй восходящей системы» или «восходящей ретикулярной активирующей системы», в отличие от «первой восходящей системы», включающей так называемые «лемнисковые» пути. Восходящая ретикулярная активирующая система представляет собой полисинаптические пути проведения в кору импульсов неспецифической природы, связанных с «пробуждением и побуждением». Белое вещество продолговатого мозга включает проводящие пути восходящего и нисходящего направлений: Нисходящие проводящие пути: • Tr. corticospinalis (pyramidalis) занимает вентральное положение, с каждой стороны от передней срединной щели, на границе со спинным мозгом подвергается неполному перекресту, который получает название перекреста пирамид или моторного перекреста, decussatio pyramidum (decussatio motoria). • Проводящие пути экстрапирамидной системы: tr. rubrospinalis, tr. tectospinalis, tr. vestibulospinalis, tr. reticulospinalis, tr. olivospinalis, fasciculus longituclinalis medialis. Восходящие проводящие пути: • Lemniscus medialis, медиальная петля, образована аксонами вторых нейронов проводящих путей проприо- и экстероцептивной чувствительности противоположной стороны. • Tr. spinothalamicus, спинно-таламический путь – проводящий путь экстероцептивной чувствительности коркового направления. В области мозгового ствола получает название спинномозговой петли, lemniscus spinalis. На всем протяжении до зрительного бугра она сопровождает волокна медиальной петли, располагаясь снаружи от проводников проприоцептивной чувствительности. • Tr. spinocerebellahs ventralis (Gowers), передний спинно-мозжечковый путь – проводящий путь проприоцептивной чувствительности. Располагается в латеральной части продолговатого мозга между оливой и нижними ножками мозжечка. Волокна заднего спинно-мозжечкового пути (Flechsig) покидают продолговатый мозг и в составе нижних ножек мозжечка направляются в кору червя.
МОСТ Мост, pons, расположен между ножками мозга сверху и спереди и продолговатым мозгом снизу и сзади. Внешнее строение. На вентральной поверхности моста находятся следующие образования: • Бульбарно-мостовая борозда, расположенная между продолговатым мозгом и нижним краем моста, где находятся корешки отводящего (VI) нерва. • Мостомозжечковый угол (треугольник), который лежит между мостом, продолговатым мозгом и мозжечком. Здесь локализуются волокна лицевого (VII)и преддверно-улиткового (VIII)нервов. • Тройнично-лицевая линия, соединяет места выхода корешков тройничного (V) и лицевого (VII) нервов и является боковой границей моста. • Средние ножки мозжечка, лежат снаружи от тройнично-лицевой линии. • Базилярная борозда, в которой лежит одноименная артерия (a. basilaris). Дорсальная часть моста принимает участие в образовании ромбовидной ямки, которая является дном IV желудочка. Здесь на границе с продолговатым мозгом определяются лицевой бугорок, colliculus facialis, и мозговые полоски IV желудочка, striae medullares ventriculi quarti, которые являются частью слухового пути. Внутреннее строение. Волокна трапециевидного тела разделяют мост на вентральную, pars basilaris, и дорсальную, pars dorsalis (tegmentum), части. В pars ventralis pontis серое вещество представлено собственными ядрами моста, nuclei proprii pontis, которые образованы телами ассоциативных нейронов проводящего пути, соединяющего кору больших полушарий с корой мозжечка (tr. cortico-ponto-cerebellaris). В pars dorsalis pontis серое вещество представлено: Ядрами черепных нервов: • тройничного (V) – двигательное и чувствительное соматические ядра; • отводящего (VI) – двигательное соматическое ядро; • лицевого (VII)– двигательное и чувствительное соматические и вегетативное парасимпатическое ядра; • преддверно-улиткового (VIII)– чувствительные ядра. При этом наиболее дорсальное положение занимает двигательное ядро отводящего нерва. Ядро лицевого нерва лежит значительно более вентрально. Волокна, выходящие из ядра лицевого нерва, направляются кверху, огибают ядро отводящего, образуя колено лицевого нерва, и снова поворачивают в вентральном направлении, выходя в мосто-мозжечковом углу. Ядра тройничного нерва располагаются снаружи и выше ядра VII пары. Чувствительные ядра преддверно-улиткового нерва находятся на границе с продолговатым мозгом снаружи от corpus trapezoideum. • серое вещество покрышки моста также образовано многочисленными ядрами ретикулярной формации ствола. Белое вещество – система восходящих и нисходящих проводящих путей. Восходящие пути составляют: • медиальная петля, lemniscus medialis; • спинномозговая петля, lemniscus spinalis; • латеральная петля, lemniscus lateralis, (является продолжением волокон трапециевидного тела и составляет часть слухового пути). Нисходящие волокна образованы проводящими путями экстрапирамидной системы (fasciculus longitudinalis medialis, tr. tectospinalis, tr. reticulospinalis и др.). Белое вещество вентральной части моста представлено продольно и поперечно идущими волокнами, fibrae longitudinales и fibrae transversae. Продольные волокна включают две составные части пирамидного пути (t r. corticospinalis и t r. corticonuclearis) и корково-мостовые волокна, которые соединяют кору лобной, затылочной и височной долей с собственными ядрами моста (fibrae corticopontinae). Поперечно идущие пучки волокон начинаются от собственных ядер моста и направляются в среднюю мозжечковую ножку противоположной стороны (fibrae ponto-cerebellares). Поперечные волокна, которые локализуются центрально от пирамидных путей, называются поверхностными, fibrae transversae superficiales, а лежащие дорсально – глубокими, fibrae transversae profundae.
МОЗЖЕЧОК Мозжечок, cerebellum, (малый мозг). Основное значение мозжечка состоит в дополнении и коррекции деятельности остальных двигательных центров. Мозжечок принимает участие в рефлекторной регуляции позы и мышечного тонуса; обеспечивает коррекцию медленных целенаправленных движении в ходе их выполнения и координацию этих движений с рефлексами поддержания позы; а также обеспечивает правильное выполнение быстрых целенаправленных движений, команда к которым поступает из коры больших полушарий. Внешнее строение. Состоит из непарной срединной части, которая называется червь, vermis, и боковых частей – полушарий мозжечка, hemispheria cerebelli. В мозжечке различают верхнюю и нижнюю поверхности, передний и задний края, на которых находятся одноименные вырезки, передние, задние и боковые углы.Посередине нижней поверхности находится широкая борозда – долинка мозжечка, valecula cerebelli, в которой помещается продолговатый мозг. Рельеф мозжечка представлен многочисленными бороздками, sulci cerebelli, которые отделяют друг от друга узкие извилины, gyri cerebelli. На границе верхней и нижней поверхностей мозжечка проходит глубокая поперечная борозда, sulcus horizontalis. Полушария разделяются постоянными глубокими бороздами на три доли: lobus anterior, lobus posterior и lobus fiocculonodularis, которым соответствуют определенные участки червя. Каждая доля разделена на дольки, которые обозначаются как классическими названиями, так и широко распространенной нумерацией по Ларселу (римские цифры). В ходе филогенеза происходит постепенное усложнение строения мозжечка параллельно развитию полушарий большого мозга. В соответствии с филогенетическим возрастом различают три части мозжечка: • старая часть, archaeocerebellum, к которой относятся клочок, узелок и язычок червя; • древняя часть, paleocerebellum, которая состоит из язычка, центральной дольки, вершины и пирамиды червя, а также крыла центральной дольки и четырехугольной дольки (передняя часть) полушарий; в филогенезе появляется после старой части; • новая часть, neocerebellum, представленная скатом, листком и бугром червя, четырехугольной (задняя часть), верхней и нижней полулунными дольками и миндалиной полушарий мозжечка; в филогенезе появляется позднее остальных частей. Эта классификация соответствует подразделению органа на отделы в соответствии с проекцией афферентных волокон мозжечкового направления. Так, старая часть называется vestibulocerebellum, так как здесь заканчиваются пути из вестибулярных ядер. Древняя часть получает название spinocerebellum, так как в нее приходят пути из спинного мозга. Наконец, новая часть, pontocerebellum, получает афферентные волокна из коры больших полушарий. Внутреннее строение. Вся наружная поверхность органа покрыта серым веществом, cortex cerebelli. Кора мозжечка имеет трехслойное строение: • Молекулярный слой – наружный слой коры. • Слой грушевидных нейронов или слой клеток Пуркинье – средний слой коры. • Зернистый слой – внутренний слой коры. Он состоит из многочисленных мелких клеток-зерен и более крупных клеток Гольджи. В толще мозжечка серое вещество представлено ядрами: • зубчатое ядро, nucleus dentatus, имеет вид изогнутой пластинки с медиально-расположенными воротами; • пробковидное ядро, nucleus emboliformis, расположено спереди от ворот зубчатого ядра; • шаровидное ядро, nucleus globosus, находится с медиальной стороны от зубчатого ядра; • ядро шатра, nucleus fastigii, – самое внутреннее из ядер мозжечка. Кора червя и полушарий мозжечка характеризуется соматотопической организацией: в передних отделах полушарий представлены верхние конечности, а в задних отделах – нижние; в передних отделах коры червя – голова и шея, а в задних отделах – туловище. Проксимальные отделы конечностей проецируются медиальнее, дистальные – латеральнее; полушария ответственны за координацию движений конечностей, червь – туловища. Белое вещество мозжечка получило название мозгового тела, corpus medullare, от которого отходят сначала более крупные мозговые пластинки белого вещества, laminae medullares, которые, в свою очередь, отдают вторичные мозговые листки. Последние дают начало еще более мелким листочкам, которые покрыты серым веществом и образуют извилины мозжечка, gyri cerebelli. Картина расположения бело
|
|||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-07-16; просмотров: 366; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.119.135.231 (0.016 с.) |