Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Роль генетического материала в дифференцировке клетокСодержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
На чем же основана клеточная дифференцировка? Развитие представлений о механизмах цитодифференцировки представлено на рис. 3.6. В конце XIX в. А. Вейсман предложил механическую модель цито-дифференцировки. Он предполагал, что при делении клеток зародыша распределение генетического материала не происходит равномерно. Только линия половых клеток получает при делении и передает потомкам полный набор хромосом. Линии же соматических клеток наследуют лишь часть генетического материала, причем все они отличаются друг от друга по количеству и содержанию полученного материала. Предположение Вейсмана нашло ряд подтверждений. Так, в ходе делений дробления у аскариды наблюдается элиминация, т.е. утрата, части хромосом. Это явление получило название диминуция хроматина. Полный набор неповрежденных (интактных) хромосом сохраняется только в том бластомере, который в дальнейшем дает начало первичным половым клеткам. Элиминация целых хромосом была обнаружена у некоторых насекомых, низших ракообразных и даже одного из представителей сумчатых млекопитающих. У последних все соматические клетки содержали лишь одну половую хромосому Х, а предшественники половых клеток - две хромосомы: ХХ или ХY в зависимости от пола животного. В дальнейшем выяснилось, что в ходе цитодифференцировки количество генетического материала может не только уменьшаться, но и увеличиваться. У некоторых насекомых, моллюсков, круглых червей в клетках слюнных желез, эпителии желудка и задней кишки, мальпигиевых сосудах и ряде других тканей обнаружены политенные хромосомы. Такие хромосомы, содержащие до тысячи и даже более копий одной и той же молекулы ДНК, образуются в результате многократной ее репликации, не сопровождающейся последующим разделением этих молекул. Явление амплификации - многократного избирательного копирования отдельных генов, наблюдаемое, в частности, в овогенезе амфибий, также приводит к увеличению количества генетического материала в определенных клетках особи (см. п. 2.4.3.4). Однако все приведенные примеры являются скорее исключением, нежели правилом. Как установлено к настоящему времени, сбалансированность генотипа по дозам генов - одно из основополагающих условий нормального развития особи. Действительно, формирование новых организмов в подавляющем большинстве случаев происходит из одной или нескольких диплоидных соматических клеток, которые делятся митозом. Этот механизм деления обеспечивает равномерное распределение генетического материала в дочерних клетках и генетическую идентичность материнской и дочерних клеток. Следовательно, все соматические клетки, образующиеся в ходе развития, среди которых и первичные половые клетки, имеют полный набор генетического материала. (Редкие случаи соматических мутаций учитывать не будем.) Наиболее прямые доказательства эквивалентности геномов соматических клеток получены методами молекулярной гибридизации нуклеиновых кислот (см. п. 5.2.2.3). Уже более 20 лет тому назад их применение показало, что ДНК всех типов клеток большинства позвоночных животных имеют одинаковое количество и одинаковые типы последовательностей нуклеотидов. Функциональная полноценность генетического материала соматических клеток была доказана в экспериментах Дж. Гердона. Суть опытов заключалась в следующем (см. рис. 3.2). Ядро яйцеклетки шпорцевой лягушки, которое содержало два ядрышка, подвергали ультрафиолетовому облучению, чем вызывали его инактивацию. Вместо него в цитоплазму яйцеклетки помещали ядро с одним ядрышком, взятое из дифференцированных клеток кожи взрослой особи или кишечника головастика. В определенном проценте случаев из такой яйцеклетки развивался нормальный организм, все клетки которого имели ядро с одним ядрышком. Полученный результат доказывает, что ядро дифференцированной клетки содержит всю информацию, необходимую для полноценного развития особи. О функциональной полноценности генетического материала зрелой соматической клетки свидетельствуют и выполненные к настоящему времени многочисленные эксперименты по клонированию растений, животных, в том числе и млекопитающих. Клонирование, в основе которого лежит трансплантация ядер дифференцированных соматических клеток в энуклеированные (лишенные ядра) ооциты, представило неопровержимые доказательства того, что геном эукариотических клеток не претерпевает необратимых изменений в ходе их дифференцировки и может быть репрограммирован, т.е. возвращен на уровень функциональной активности, наблюдаемый у зиготы. Более того, показано, что ядра высокодифференцированных клеток, таких как В- или Т-лимфоциты, способны обеспечить полноценное развитие особи в сходных экспериментах, несмотря на то что некоторые их гены (иммуноглобулинов и Т-рецепторов) претерпевают перестройку в ходе дифференцировки. И хотя остается неясным, способны ли сохранять возможность реализации информации гены любых типов дифференцированных клеток, список обладающих такой способностью клеток достаточно велик и включает у млекопитающих в том числе фибробласты эмбрионов и взрослых животных, эпителиальные клетки молочной железы и яйцевода, В- и Т-лимфоциты, незрелые клетки Сертоли и пролиферирующие нейральные клетки коры головного мозга эмбрионов. Следовательно, причина дифференцировки - не изменение количества или функциональной полноценности генетического материала в клетках, а какие-то иные механизмы. В настоящее время считают, что возникающая в процессе развития специализация клеток - результат дифференциальной (или избирательной) экспрессии генов. Эта точка зрения ведет начало от Т. Моргана, который, опираясь на хромосомную теорию наследственности, предположил, что дифференцировка клеток в процессе онтогенеза является результатом последовательных реципрокных (взаимных) влияний цитоплазмы и меняющихся продуктов активности генов. Различные типы клеток используют разные гены из одинакового набора, присутствующего в каждой клетке. Это означает, что в конкретных клетках активны не все гены, а только часть из них, причем экспрессия тех или иных генов происходит избирательно в зависимости от типа клеток, этапа онтогенеза и других факторов. Результатом такой избирательной экспрессии становится образование в разных типах клеток различных наборов белков, которые обеспечивают протекание в клетках определенных биохимических реакций, специфичность их строения и функции. Так, нервные клетки способны возбуждаться и передавать это возбуждение на другие клетки, эритроциты - транспортировать кислород к тканям, мышечные клетки - сокращаться и тем самым обеспечивать различные проявления движения, фоторецепторы - воспринимать световой поток. Выполнение этими клетками специфических функций определяется их строением, а именно: наличием отростков у нейронов, по которым передается возбуждение; двояковогнутой формой эритроцитов, позволяющей им проникать в узкие капилляры и осуществлять газообмен; значительной протяженностью мышечных волокон, образованных при слиянии нескольких клеток-предшественников, что делает их способными эффективно изменять свою длину; формированием складок мембраны, где располагается фотопигмент, у палочек и колбочек. Указанные морфофункциональные различия обеспечиваются разнообразием белков: нейроны продуцируют нейропептиды, эритроциты - гемоглобин, мышечные клетки - актин и миозин, клетки сетчатки - опсины. В некоторых случаях дифференцировка оказывается связанной с синтезом не белков, а других веществ, например сахаров и их производных. Так, межклеточное вещество хрящевой ткани состоит из мукополисахаридов - производных углеводов. Однако их синтез в клетках-хондробластах невозможен без некоторых специфических ферментов, а последние - это белки. Поэтому утверждение, что в основе подавляющего большинства клеточных дифференцировок лежит синтез специфических белков, абсолютно справедливо. Следует подчеркнуть, что этот принцип диффе-ренцировки является общим в онтогенезе как животных, так и растений, несмотря на то что между ними существуют огромная эволюционная дистанция и существенные различия в характере развития. Гипотезу избирательной активности генов подтверждают данные, полученные в генетических и эмбриологических исследованиях. Один из объектов, позволяющих визуально, с помощью светового или электронного микроскопа изучать активность отдельных генов, - политенные хромосомы. В них хорошо видны темноокрашенные области плотной упаковки ДНК, которые получили название дисков. Между ними расположены светлые участки генетического материала менее плотной упаковки. Многие отдельные диски соответствуют отдельным генам. Активная транскрипция определенных генов в таких хромосомах сопровождается образованием вздутий или пуфов на месте дисков (рис. 8.21). Сравнение различных типов дифференцированных клеток по набору только что транскрибированных молекул пре-мРНК, выполненное методом молекулярной гибридизации молекул РНК с комплементарными им ДНК, также выявило различия в зависимости от типа клеток. Было показано, что клетки разных типов, синтезирующие отличающиеся мРНК и белки, демонстрируют в одних и тех же хромосомах различную локализацию пуфов. Кроме того, расположение пуфов меняется в ходе онтогенетического развития, что коррелирует с синтезом определенных белков в конкретный промежуток времени (рис. 8.21).
Рис. 8.21. Последовательное изменение активности трех пуфов с особой морфологией - колец Бальбиани (КБ1-КБЗ) у Chironomus tentans: 1 - диск; 2 - междисковый промежуток; 3 – пуф
По мере развития число функционирующих генов в клетке, по-видимому, прогрессивно снижается. Так, из 40 тыс. генов одного из видов морского ежа на стадии бластулы активны примерно 30 тыс., гаструлы и личинки - 12-15, у взрослых животных - 5-7 тыс. генов. У человека, по некоторым оценкам, в раннем эмбриогенезе активны до 60% генов, а в дифференцированных клетках взрослого организма - от 1 до 7-10% (по отдельным данным, до 44% в нервных клетках). Установлено, что часть генов при усилении специализации блокируется в клетках необратимо. Подтверждением тому могут служить уже упоминавшиеся опыты Гердона по пересадке ядра дифференцированной клетки в яйцеклетку с инактивированным собственным ядром. Количество успешных развитий особей прямо зависело от возраста донора (рис. 8.22). С этой точки зрения дифференцировка может быть определена как экспрессия той части генома, которая остается в распоряжении клетки. Вместе с тем эти опыты обнаружили и другие важнейшие закономерности. Так, они еще раз подтвердили предположение Т. Моргана о решающем значении взаимодействия цитоплазмы и ядра в жизнедеятельности клеток и развитии организма. Как было сказано ранее (см. п. 2.4.4.4-е), структурные гены генотипа подразделяются на две группы: гены «домашнего хозяйства», кодирующие белки, обеспечивающие реализацию фундаментальных процессов жизнедеятельности в клетке, и гены дифференцировки, называемые также гены «роскоши». Последние отвечают за синтез специфических белков. На самых ранних этапах эмбрионального развития, а именно в ходе дробления, когда начинает работать собственный геном зародыша, первоначально экспрессируются только гены «домашнего хозяйства». Синтез белков, кодируемых генами «роскоши», в клетках большинства
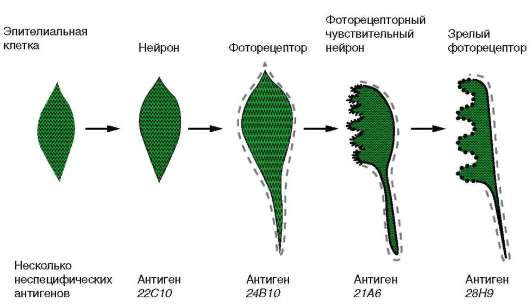
Рис. 8.22. Зависимость успеха пересадки ядер из дифференцированной клетки в яйцеклетку от возраста донора (i-VI) ядра: I - бластула; II - гаструла; III - нейрула; IV - появление мышечной реакции; V - начало сердечной деятельности и вылупления; VI - активное плавание; 1 - ранняя гаструла; 2 - нейрула; 3 - плавающий головастик; 4 - питающийся головастик; вверху изображена схема опыта хордовых осуществляется, начиная со стадии бластулы. В этой группе представлены гены, кодирующие тканеспецифические белки, характерные для всех типов клеток данной ткани (например, нервной), и гены, кодирующие типоспецифические белки, определяемые только в конкретных специализированных клетках (например, в колбочках). Первоначально на этапах дифференцировки включаются гены, отвечающие за синтез тканеспецифических белков, а затем - типоспецифических. Проследить это возможно на примере синтеза белков мембраны. Клетки разных типов характеризуются различными белковыми компонентами клеточной поверхности, которые к тому же изменяются по мере развития клеток. Эти специфические мембранные белки часто выявляют с помощью антисывороток, поэтому их называют антигенами диф-ференцировки. На рис. 8.23 показаны временные изменения клеточной мембраны одной эпителиальной клетки дрозофилы по мере того, как она превращается в фоторецептор сетчатки. Как только эпителиальная клетка приобретает свойства нейрона, она экспрессирует антиген 22С10, который обнаруживается и на других нервных клетках. Вскоре клетка начинает синтезировать другую молекулу клеточной мембраны - антиген 24В10, что характерно только для нейронов, дающих начало фоторецепторам. На более поздних стадиях в некоторых областях созревающего фоторецептора появляется антиген 21А6, а затем другой, 28Н9, специфические для окончательно дифференцированных фоторецепторов сетчатки.
Рис. 8.23. Избирательность синтеза специфических белков клеточной мембраны в ходе дифференцировки нейрона в фоторецептор сетчатки
Число активных генов терминальной дифференцировки в специализированных клетках очень невелико. При изучении разнообразия мРНК в почках, печени и головном мозге мышей было обнаружено, что большая их часть была одинакова и представляла собой результат транскрипции генов «домашнего хозяйства». Лишь примерно 1/10 из общего количества активных генов, число которых, как было сказано выше, составляет 1-10%, оказались специфичны для какой-либо одной ткани (т.е. всего около 0,1-1% от общего числа генов генома). Именно они транскрибировались с уникальных нуклеотидных последовательностей генов «роскоши». Часть клеток развивающегося организма не вступает на путь дифференцировки, сохраняя способность к самообновлению и потенциал к развитию, - стволовые клетки (см. п. 3.1.2). Во взрослом организме присутствуют региональные стволовые клетки, которые могут дать начало одному, нескольким или многим типам специализированных клеток. Они найдены не только в хорошо регенерирующих в норме тканях, таких как эпителий и красный костный мозг, но также и в статических, образующих, например, нервную систему и печень. Стволовые клетки играют центральную роль в гистогенезе и, кроме того, составляют существенный восстановительный резерв в организме, способствуя замещению дефектов, возникающих в силу тех или иных обстоятельств в разных органах. Избирательная экспрессия генов, наблюдаемая в ходе клеточной дифференцировки, основана на действии целого ряда механизмов, которые условно можно разделить на две группы: локальные - внутриклеточные, обеспечивающие избирательную экспрессию генов в отдельной клетке, и системные (межклеточные взаимодействия, эмбриональная индукция, нервная и гуморальная регуляция). Последние, определяя целостность развивающегося организма и достижение определенного конечного результата, регулируют дифференцировку клеток в строго определенном направлении и закономерное расположение различных дифференцированных клеточных типов в целом организме. Еще одним важным механизмом, обеспечивающим процесс дифференцировки и целостное развитие организма, является гетерогенность яйцеклетки (овоплазматическая сегрегация). Рассмотрим различные механизмы регуляции экспрессии генов более подробно.
|
||||
|
|
Последнее изменение этой страницы: 2016-07-15; просмотров: 471; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.149.29.209 (0.011 с.) |