Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Тема 1. Фізіологічні механізми м’язевого скороченняСодержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
Тема 1. ФІЗІОЛОГІЧНІ МЕХАНІЗМИ М’ЯЗЕВОГО СКОРОЧЕННЯ СТРУКТУРА ТА ФУНКЦІЇ СКЕЛЕТНОГО М’ЯЗУ Скелетний м’яз являє собою складно побудований орган. Скоротлива функція м’язу здійснюється завдяки діяльності м’язових скорочень, які володіють властивістю збудливості і скоротливості; вони складають значну частину маси м’язу. М’язові волокна, що утворюють скелетні м’язи, відносяться до поперечнопосмугованої м’язової тканини. Зовнішня оболонка м’язу побудована сполучною тканиною, і має назву епімізій. Він охоплює м'яз і надає йому форму. М’яз складається з пучків м’язових волокон, що вкриті також сполучною тканиною. Сполучнотканинна оболонка, що охоплює кожний пучок, називається перимізієм. Під перимізієм розташовуються м'язові волокна, що є окремими м'язовими клітинами. Кожне м'язове волокно також покриває сполучнотканинна оболонка, котра називається ендомізієм. М'ЯЗОВЕ ВОЛОКНО Діаметр м'язових волокон коливається від 10 до 80 мікрометрів (мкм) і вони практично невидимі неозброєним оком. Більшість із них тягнеться на усю довжину м'яза. Наприклад, довжина м'язового волокна стегна може перевищувати 35 см. Кількість волокон у м'язі значно коливається залежно від його розміру та функції. Сарколема. Якщо уважно розглядати окреме м'язове волокно, то можна помітити, що воно вкрите плазматичною мембраною — сарколемою. На кінці кожного м'язового волокна його сарколема з'єднується з сухожилком, прикріпленим до кістки. Саркоплазма. За допомогою мікроскопа можна побачити, що всередині сарколеми м'язове волокно містить послідовно дрібніші субодиниці (рис. 1). Найбільші з них — міофібрили, які являють собою паличкоподібні структури, котрі тягнуться на всю довжину волокон. Простір між ними заповнений желатиноподібною рідиною. Це - саркоплазма, рідинна частина м'язового волокна, що є його цитоплазмою. Саркоплазма містить головним чином розчинні білки, мікроелементи, глікоген, жири та необхідні органели. Вона відрізняється від цитоплазми тим, що більшість її клітин містить більшу кількість накопиченого глікогену, а також киснесполучною сполукою — міоглобином, подібним до гемоглобіну. Поперечні трубочки. Саркоплазму перетинає широка сітка поперечних трубочок (Т-трубочок), котрі є продовженням сарколеми (плазматичної мембрани). Вони взаємоз'єднуються, проходячи серед міофібрил, і забезпечують швидку передачу нервових імпульсів, котрі сприймає сарколема, до окремих міофібрил. Окрім того, трубочки забезпечують надходження до внутрішніх частин м'язового волокна речовин, котрі переносяться позаклітинними рідинами: глюкози, кисню, різних іонів. Саркоплазматичний ретикулум. У м'язовому волокні також міститься сітка поздовжніх трубочок, котра називається саркоплазматнчним ретикулумом (СР). Ці мембранні канальця розташовані паралельно міофібрилам, утворюючи петлі навколо них. СР слугує місцем накопичення кальцію, котрий має велике значення для м'язового скорочення.
Рис. 1. Будова (а) та поперечиний розріз (б) скелетного м'яза
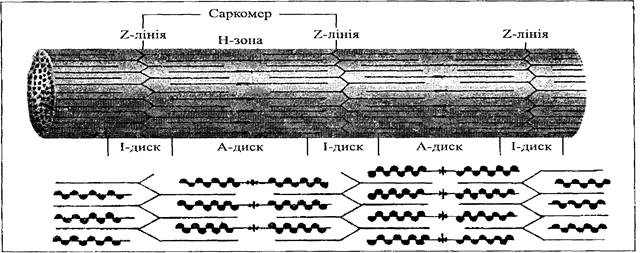
МІОФІБРИЛА У кожному м'язовому волокні міститься від кількох сотен до кількох тисяч міофібрил. Це скоротливі елементи скелетного м'яза. Міофібрила складаються з саркомерів, що мають вигляд довгих ниток. Смуги та саркомір. Волокна скелетного м'яза під мікроскопом мають характерний смугастий вигляд. Саме тому скелетний м'яз називають також поперечносмугастим. Такі ж смуги є характерними для серцевого м'яза, тому його також можна вважати поперечносмугастим. На рис. 2, що демонструє будову міофібрили, чітко видно смуги. Видно те, що темні ділянки, так звані А-диски, чергуються зі світлими, котрі називаються І-дисками. У центрі кожного А-диска є світліша ділянка — Н-зона, котру можна побачити лише при розслабленні міофібрили. Світлі І-диски перетинаються темною Z-лінією. Основною функціональною одиницею міофібрили є саркомір. Кожна міофібрила складається з численної кількості саркомерів, кінці котрих з'єднані один з одним біля Z-ліній. Кожний саркомір включає те, що знаходиться між кожною парою Z-ліній, у такій послідовності:
• І-диск (світла ділянка); • А-диск (темна ділянка); • Н-зона (у середині А-диска); • решта А-диска • другий І-диск.
Рис.2. Будова міофібрил
Рис. 3. Основна функціональна одиниця міофібрили — саркомір, котрий складається із певним чином розташованих актиновнх та міозннових філаментів
Тонкіші філаменти, що відповідають за мязове скорочення утворені актином, грубіші — міозином. У кожній міофібрилі порядрозташовуються близько 3000 актинових та 1500 міозинових філаментів. Смуги, характерні для м'язових волокон, є результатом розташування цих філаментів, що ілюструє рис. 3. Світлий І-диск вказує на ділянку саркоміра з тонкими актиновими філаментами. Темний А-диск являє собою ділянку, що складається як з грубих міозинових, так і з тонких актинових філаментів. Н-зона — центральна частина А-диска, що є видимою тільки тоді, коли саркомір перебуває у спокої. Вона складається виключно з грубих філаментів. Світліше забарвлення Н-зони порівняно з сусіднім А-диском обумовлене відсутністю у ній актинових філаментів. Н-зону можна побачити тільки коли саркомір розслаблений, оскільки він вкорочується при скороченні й актинові філаменти «втягуються» у цю зону, роблячи її забарвлення таким, як і решта частин А-диска. Міозинові філаменти. Хоча і було відмічено, що у кожній міофібрилі міститься близько 3000 актинових та 1500 міозинових філаментів, ці цифри є не зовсім точними. Близько 2/3 білків скелетного м'яза становить міозин. Міозинові філаменти грубі (товсті). Кожний міозиновий філамент утворений приблизно 200 молекулами міозину, розташованими поряд кінцями один до одного. Кожна молекула міозину складається з двох сплетених протеїнових пучків. Один кінець кожного пучка утворює, глобулярну голівку, так звану міозинову голівку. Кожний філамент має кілька таких голівок, котрі випинаються вперед і утворюють поперечні містки, що взаємодіють під час м'язового скорочення зі спеціальними активними ділянками на актинових філаментах. Актинові філаменти. Один кінець кожного актинового філамента входить до Z-лінії, другий протягується до центра саркомера, проходячи між міозиновими філаментами. Кожний актиновий філамент має активну ділянку, до котрої може «прив'язатися» міозинова голівка. Кожний тонкий, або актиновий, філамент складається з трьох різних протеїнових молекул: актину, тропоміозину та тропоніну. Актин утворює основу філамента. Окремі актинові молекули є глобулярними і, з'єднуючись разом, утворюють нитки актинових молекул. Дві нитки обвиті одна навколо другої у вигляді спіралі подібні до двох низок перлин, скручених разом. Тропоміозин — це білок, що має форму трубки; він обвиває актинові нитки, заповнюючи заглиблення між ними. Тропонін є складнішим білком, котрий через рівні проміжки прикріплений до ниток актину та до тропоміозину. Складна діяльність тропоміозину та іонів кальцію спрямована на збереження стану розслаблення або розвиток скорочення міофібрили, про які йтиметься далі. Джерела енергії Продукти харчування складаються, в основному, з вуглецю, водню, кисню, а при наявності білків — з азоту. Молекулярні зв'язки у харчових продуктах відносно слабкіші, тому при розщепленні вивільнюється невелика кількість енергії. Отже, продукти харчування не використовуються безпосередньо для потреб клітини. Енергія молекулярних зв'язків продуктів харчування хімічно вивільнюється у клітинах організму й зберігається у вигляді високоенергетичного сполучення — аденозинтрифосфату (АТФ). Утворення АТФ дозволяє клітинам зберігати енергію у цьому високоенергетичному сполученні. У спокої енергія, необхідна нашому організму, забезпечується за рахунок розщеплення практично однакової кількості вуглеводів та жирів. Білки являють собою «будівельні блоки» і звичайно забезпечують функціонування клітин невеликою кількістю енергії. При збільшенні м'язового зусилля у якості джерела енергії більше використовуються вуглеводи. При максимальному короткочасному навантаженні АТФ майже виключно утворюється за рахунок вуглеводів. Вуглеводи. Залежність м’язів від вуглеводів під час фізичного навантаження пов'язана з їх наявністю, а також здатністю м'язової системи їх розщеплювати. Вуглеводи насамкінець перетворюються на глюкозу — моносахарид, котрий транспортується кров'ю до усіх тканин організму. У стані спокою ввібрані вуглеводи попадають у м'язи та печінку, а потім перетворюються на більш складну молекулу цукру — глікоген. Глікоген знаходиться у цитоплазмі допоки клітини не використають його для утворення АТФ. Глікоген, що міститься у печінці, може зновуперетворюватися на глюкозу. Він транспортується кров'ю до активних тканин, де й відбувається його метаболізм (розщеплення). Таблиця 1. Запаси палива та енергії в організмі
Примітка. Оцінку зроблено на основі середньої маси тіла 65 кг з вмістом жиру 12 %.
Вміст вуглеводів у печінці та скелетних м'язах обмежений; їх вистачає для утворення не більше 2000 ккал енергії. Ця кількість витрачається на те, щоб пробігти 32 км. Запаси жирів достатні для утворення понад 70000 ккал енергії. Вміст глікогену у печінці та м'язах обмежений, його запаси можуть вичерпатися, якщо у раціоні харчування немає достатньої кількості вуглеводів. Таким чином, поповнення запасу багато у чому залежить від харчових джерел крохмалів та цукру. Без достатнього споживання вуглеводів м'язи і печінка не мають свого основного джерела енергії. Жири й білки також використовуються як джерела енергії. В організмі міститься значно більше жирів, ніж вуглеводів. Однак жири є менш доступними клітинному метаболізму, оскільки, перш за все, має бути розщеплена складна форма — тригліцерид — на основні компоненти: гліцерин та вільні жирні кислоти. Тільки вільні жирні кислоти використовуються для утворення АТФ. З однакової кількості жирів та вуглеводів утворюється абсолютно різна кількість енергії, відповідно 9 та 4 ккал. У будь-якому випадку інтенсивність вивільнення енергії з цих сполучень дуже невелика, щоб задовольнити потреби організму в енергії під час інтенсивної м'язової діяльності. Білки. Процес перетворення білків або жирів на глюкозу називається глюконеогенезом. У результаті серії реакцій білок може перетворитися на жирні кислоти. Це називається ліпогенез. Білки забезпечують 5-10 % енергії, необхідної для виконання тривалої фізичної вправи. Для утворення енергії використовуються лише основні одиниці білка — амінокислоти. Щоб бути корисною, енергія має вивільнятися з хімічних сполук з контрольованою інтенсивністю. Частково ця інтенсивність визначається вибором джерела енергії. Якщо використовується велика кількість енергії з одного джерела, клітини розраховують, головним чином, саме на це джерело. Такий вплив наявності енергії називається ефектом масового впливу. Спеціальні ферменти забезпечують чіткіший контроль інтенсивності вивільнення енергії. Багато з них полегшують розщеплення (катаболізм) хімічних сполучень. Хоча назви ферментів досить складні, всі вони закінчуються суфіксом -аза. Наприклад, фермент, що впливає на АТФ, називається аденозинтрифосфатаза (АТФаза). УТВОРЕННЯ АТФ Молекула АТФ складається з аденозину (молекули аденіну, з'єднаної з молекулою рибози), з'єднаного з трьома групами неорганічного фосфату (Рн). При впливі ферменту АТФази остання фосфатна група відщеплюється від молекули АТФ, швидко вивільнюючи велику кількість енергії (7,6 ккал/моль АТФ). У результаті АТФ розщеплюється на АДФ (аденозиндифосфат) та фосфор. Клітини утворюють АТФ за допомогою трьох систем: системи АТФ-КФ, гліколітичної та окиснювальної. СИСТЕМА АТФ-КФ Найпростішою енергетичною системою є система АТФ-КФ. Окрім АТФ, клітини містять ще одну багату енергією фосфатну молекулу — креатинфосфат (КФ). Енергія, вивільнювана при розщепленні КФ, на відміну від енергії, що вивільнюється при розщепленні АТФ, не використовується безпосередньо для виконання роботи на клітинному рівні. Вона використовується для ресинтезу АТФ, щоб забезпечити його відносно постійне утворення. Вивільненню енергії при розщеплені КФ сприяє фермент креатинкіназа, котрий діє на КФ для відокремлення фосфору від креатину. Вивільнена енергія може бути використана для приєднання Рн до молекули АДФ. При використанні цієї системи (енергія вивільнюється з АТФ в результаті відщеплення фосфатної групи) клітини можуть запобігти вичерпаю запасів АТФ, розщеплюючи КФ і тим самим забезпечуючи енергію для утворення великої кількості АТФ. Це швидкий процес, котрий може здійснюватися без допомоги будь-яких спеціальних структур клітини. Він може відбуватися й за участю кисню, однак для його здійснення кисень не потрібний, тому систему АТФ-КФ називають анаеробною. У перші секунди інтенсивної м'язової діяльності кількість АТФ підтримується на відносно постійному рівні, тоді як рівень КФ невпинно знижується, оскільки він використовується для поповнення запасів АТФ. У стані виснаження рівні АТФ та КФ є досить низькими і не можуть забезпечити енергію для наступних скорочень та розслаблень м'язів. Таким чином, підтримання рівня АТФ за рахунок енергії, що вивільнюється при розщепленні КФ, є обмеженим. Запаси АТФ та КФ є достатніми для задоволення енергетичних потреб м'язів лише протягом 3-15с спринтерського бігу. Після цього м'язам доводиться розраховувати на інші процеси утворення АТФ: гліколітичний та окиснювальний. ГЛІКОЛІТИЧНА СИСТЕМА Інше джерело отримання АТФ передбачає вивільнення енергії в результаті розщеплення (лізису) глюкози. Це — гліколітична система, котра включає процес гліколізу, тобто розщеплення глюкози за допомогою спеціальних гліколітичних ферментів. Глюкоза становить близько 99 % усіх цукрів, що циркулюють у крові. Вона надходить у кров в результаті засвоєння вуглеводів та розщеплення глікогену печінки. Глікоген синтезується з глюкози внаслідок процесу, що називається глікогенезом. Глікоген міститься у печінці або м'язах, доки не стане потрібним. Перш ніж глюкоза або глікоген можуть бути використані для утворення енергії, вони мають трансформуватися у сполучення, котре називається глюкозо-6-фосфат. Для перетворення молекули глюкози необхідна одна молекула АТФ. При розщепленні глікогену глюкозо-6-фосфат утворюється з глюкозо-1-фосфату без витрати енергії. Гліколіз починається, як тільки утворюється глюкозо-6-фосфат. Закінчується гліколіз утворенням піровиноградної кислоти. Коли ми говоримо про гліколітичну систему, ми маємо на увазі, що процес гліколізу перебігає без участі кисню. У цьому випадку піровиноградна кислота перетворюється на молочну кислоту. Гліколіз, що є складнішим процесом, ніж система АТФ-КФ, забезпечує розщеплення глікогену на молочну кислоту завдяки 12 ферментним реакціям. Усі ці ферменти знаходяться у цитоплазмі клітин. У результаті гліколізу утворюється 3 молі АТФ на кожний моль розщепленого глікогену. Якщо замість глікогену використовується глюкоза, то утворюється усього 2 молі АТФ, оскільки 1 моль витрачається на перетворення глюкози на глюкозо-6-фосфат. Ця енергетична система не забезпечує утворення великої кількості АТФ. Незважаючи на це, сукупні дії гліколітичної системи та системи АТФ-КФ забезпечують продукування сили м'язами навіть при обмеженому надходженні кисню. Ці дві системи домінують у перші хвилини виконання вправ високої інтенсивності. Іншим значним недоліком анаеробного гліколізу є те, що він викликає накопичення молочної кислоти у м'язах та рідинах організму. У спринтерських дисциплінах тривалістю 1-2 хв потреби гліколітичної системи є дуже великими, і рівні вмісту молочної кислоти можуть збільшитися з 1 (показник у стані спокою) до понад 25 ммоль/кг. Таке підкислення м'язових волокон гальмує подальше розщеплення глікогену, оскільки порушує функцію гліколітичних ферментів. Окрім того, кислота знижує здатність волокон зв'язувати кальцій і це може перешкодити скороченню м'язів. Інтенсивність енерговитрат м'язового волокна під час навантаження може бути у 200 разів вищою, ніж у стані спокою. Гліколітична система та система АТФ-КФ не в змозі тривало забезпечити необхідну кількість енергії. Молочна кислота і лактат — не одне й те ж сполучення. Молочна кислота має формулу С3Н6О3. Лактат являє собою будь-яку сіль молочної кислоти.
ОКИСНЮВАЛЬНА СИСТЕМА Останньою системою утворення енергії клітиною є окиснювальна система, найскладніша з трьох енергетичних систем. Процес, в результаті котрого організм для продукування енергії дисимілює сполучення, багаті на енергію, за допомогою кисню, називається клітинним диханням. Це аеробний процес, оскільки у ньому бере участь кисень. АТФ утворюється у спеціальних клітинних органелах – мітохондріях. У м'язах вони примикають до міофібрил, а також розкидані по саркоплазмі. М'язи мають постійно забезпечуватися енергією для продукування сили під час тривалої м'язової діяльності. На відміну від анаеробного утворення АТФ, окиснювальна система продукує значну кількість енергії, тому аеробний метаболізм є основним методом утворення енергії під час м'язової діяльності, що потребує виявлення витривалості. Це ставить підвищені вимоги до системи транспорту кисню до активних м'язів. Окиснення вуглеводів. Окиснювальне утворення АТФ включає три процеси: 1) гліколіз; 2) цикл Кребса; 3) ланцюжок переносу електронів. Гліколіз при обміні вуглеводів відіграє важливу роль як в анаеробному, так і в аеробному утворенні АТФ. Причому він перебігає однаково, незалежно від того, чи бере участь у цьому процесі кисень. Участь кисню визначає лише «частку» кінцевого продукту — піровиноградної кислоти. При анаеробному гліколізі утворюється молочна кислота і усього 3 молі АТФ на 1 моль глікогену. За участю кисню піровиноградна кислота перетворюється на сполучення, котре називається ацетилкофермент А (ацетил-КоА). Цикл Кребса. Після утворення ацетил-КоА попадає у цикл Кребса (цикл лимонної кислоти) — складну послідовність хімічних реакцій, котрі дозволяють завершити окиснення ацетил-КоА. Наприкінці циклу Кребса утворюється 2 молі АТФ, а речовина (сполучення, на котре впливають ферменти, у цьому випадку первісний вуглевод) розщеплюється і, з'єднуючись з киснем, утворює діоксид вуглецю (СО2), котрий легко дифундує з клітин, транспортується кров'ю у легені й виділяється у зовнішнє середовище (відбувається реакція декарбоксилювання К-Н + СО2). Утворення енергії за рахунок вуглеводів. Окиснювальна система утворення енергії забезпечує отримання 39 молекул АТФ з однієї молекули глюкози. Якщо процес починається з глюкози, то утворюється 38 молекул АТФ (згадаймо, що одна молекула АТФ використовується до початку гліколізу для утворення глюкозо-6-фосфату). Окиснення жирів. Як уже відмічалося, жири також роблять свій внесок в енергетичні потреби м'язів. Запаси глікогену у м'язах та печінці можуть забезпечити усього 1200-2000 ккал енергії, у той час як жири, що містяться всередині м'язових волокон та в жирових клітинах, — близько 70 000-75 000 ккал. Хоча жирами називають багато хімічних сполук, такі, як тригліцериди, фосфоліпіди та холестерин, тільки тригліцериди використовуються як основні джерела енергії. Тригліцериди знаходяться у жирових клітинах та волокнах скелетних м'язів. Щоб використати тригліцериди для утворення енергії, необхідно розщепити їх на основні складові: одну молекулу гліцерину та три молекули вільних жирних кислот. Цей процес називається ліполізом і здійснюється ферментами — ліпазами. Вивільнившись з тригліцериду (жиру), вільні жирні кислоти можуть попасти у кров, котра транспортує їх по усьому організму, і у результаті дифузії проникнути у м'язові волокна. Інтенсивність надходження вільних жирних кислот у м'язові волокна залежить від градієнта концентрації. Підвищення концентрації вільних жирних кислот у крові виштовхує їх у м'язові волокна. Процес β-окиснення. Незважаючи на значні структурні відмінності між різними вільними жирними кислотами, їх метаболізм майже однаковий, як видно. До того, як вони попадуть у м'язові волокна, вільні жирні кислоти активуються енергією АТФ за допомогою ферментів. Таким чином їх готують до катаболізму (розщеплення) у мітохондріях. Цей ферментний катаболізм жирів мітохондріями називається β-окисненням. У цьому процесі вуглецевий ланцюжок вільної жирної кислоти ділиться на двовуглецеві рештки оцтової кислоти. Наприклад, якщо первісно вільна жирна кислота мала 16-вуглецевий ланцюжок, то при β--окисненні утворюється 8 молекул оцтової кислоти. Вся оцтова кислота потім перетворюється на ацетил-КоА. Як і при обміні глюкози, проміжними продуктами окиснення вільних жирних кислот є АТФ, Н2О, СО2. Однак для повного спалювання молекули вільних жирних кислот потребується більше кисню. Хоча жири забезпечують більше кілокалорій енергії на грам, ніж вуглеводи, для їх окиснення потребується більше кисню, ніж для окиснення вуглеводів. Жири утворюють 5,6 молекули АТФ відносно молекули О2, вуглеводи — 6,3 молекули АТФ відносно молекули О2. Доставка кисню обмежена кисневотранспортною системою, тому кращим джерелом енергії під час виконання фізичної вправи високої інтенсивності є вуглеводи. МЕТАБОЛІЗМ БІЛКІВ Як уже відмічалося, вуглеводи та жири є переважними джерелами енергії нашого організму. Однак використовуються і білки або, скоріше, амінокислоти, з котрих вони складаються. Деякі амінокислоти можуть перетворюватися на глюкозу (за допомогою глюконеогенезу). Інші можуть перетворюватися на різні проміжні продукти окиснювального метаболізму (такі, як піровиноградна кислота або ацетил-КоА), щоб взяти участь в окиснювальному процесі. Кількість енергії, утворюваної білками, досить важко визначити, на відміну від енергії, утворюваної вуглеводами або жирами, оскільки білки також містять азот. При катаболізмі амінокислот певна кількість азоту використовується для утворення нових амінокислот, решта азоту перетворюється на сечовину і виділяється головним чином з сечею. Цей процес потребує використання АТФ і, отже, призводить до витрат якоїсь кількості енергії. ОКИСНЮВАЛЬНІ ЗДАТНОСТІ М'ЯЗІВ Окиснювальна здатність м'яза - це показник її максимальної здатності використовувати кисень. Активність ферментів. Здатність м'язових волокон окиснювати вуглеводи та жири досить важко визначити. У багатьох дослідженнях спостерігали тісний взаємозв'язок між здатністю м'яза виконувати аеробну вправу протягом тривалого часу та активністю її окиснювальних ферментів. Оскільки для окиснювання потребується багато ферментів, то їх активність у м'язових волокнах є достатньо надійним показником окиснювального потенціалу. Склад м'язових волокон та тренувальні навантаження, спрямовані на розвиток витривалості. Склад волокон м'яза частково визначає його окиснювальну здатність. ПС волокна мають більшу схильність до аеробної діяльності, ніж ШС, оскільки містять більше мітохондрій та окиснювальних ферментів. ШС волокна більш придатні для гліколітичного продукування енергії. Отже, чим більше у м'язах ПС волокон, тим вища їх окиснювальна здатність. Наприклад, у найсильніших бігунів на довгі дистанції значно більше ПС волокон, мітохондрій і вища активність окиснювальних ферментів, ніж у нетренованих людей. Тренувальні навантаження, спрямовані на розвиток витривалості збільшують окиснювальні здатностіусіх волокон і особливо ШС, ставлячи високі вимоги до окиснювального фосфорилювання, вони стимулюють м'язові волокна до утворення великої кількості мітохондрій, містять велике число окислювальних ферментів. Збільшуючи кількість ферментів у волокнах для β-окиснення, такі навантаження також допомагають м'язам більшою мірою розраховувати на жири як джерело продукування АТФ. Таким чином, тренувальні навантаження на розвиток витривалості дають змогу підвищити аеробні здатності м'язів навіть у людей з високим вмістом ШС волокон. Разом з тим відомо, що ШС волокно у результаті тренування на розвиток витривалості не зможе такою ж мірою збільшити витривалість, як ПС волокно. Потреба у кисні. Хоча окиснювальна здатність м'язів визначається кількістю мітохондрій та окиснювальних ферментів у них, окиснювальний метаболізм насамкінець залежить від їх адекватного постачання киснем. У стані спокою потреби організму в АТФ відносно невеликі, тому потреба у кисні також мінімальна. Однак зі збільшенням інтенсивності навантаження зростає і потреба в енергії. Для її задоволення необхідно збільшити окиснювальне утворення АТФ. Задоволення потреб м'язів у кисні здійснюється за рахунок збільшення частоти та глибини дихання, поліпшення процесу газообміну у легенях. Серце починає скорочуватися частіше, постачаючи у м'язи більшу кількість оксигенованої крові. В організмі людини кисню небагато. Тому кількість кисню, що попадає у кров, котра проходить через легені, прямо пропорційна кількості, використовуваній тканинами для окиснювального фосфорилювання. Отже, можна досить точно визначити величину аеробного продукування енергії, вимірявши кількість кисню, що споживається у легенях. Тема 1. ФІЗІОЛОГІЧНІ МЕХАНІЗМИ М’ЯЗЕВОГО СКОРОЧЕННЯ
|
|||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-06-26; просмотров: 522; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.188.154.238 (0.014 с.) |