
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Гемоглобиновая буферная система.Содержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
Самая мощная буферная система. На ее долю приходится 75% всей буферной емкости крови. Участие гемоглобина в регуляции рН крови связано с его ролью в транспорте кислорода и углекислоты. Состав гемоглобинового буфера можно выразить так:
Гемоглобиновая буферная система состоит из HHb гемоглобина, который является слабой кислотой и его калиевой соли - Системы гемоглобина и оксигемоглобина взаимопревращающиеся системы и работают как единое целое. При насыщении гемоглобина кислородом гемоглобин становится более сильной кислотой – HНbО2 и увеличивает отдачу в раствор ионов водорода. В капиллярах легких, гемоглобин присоединяет кислород и превращается в оксигемоглобин: HHb + О2 Образование оксигемоглобина приводит к некоторому подкислению крови, вытеснению части угольной кислоты из бикарбонатов: HНbО2 + КНСО3 Щелочной резерв крови понижается. В периферических капиллярах большого круга кровообращения калиевая соль оксигемоглобина диссоциирует и гемоглобин эритроцитов отдает кислород тканям: КНbО2 В результате способность гемоглобина связывать ионы водорода увеличивается. Одновременно в эритроциты поступает углекислый газ. Под влиянием фермента карбангидразы углекислый газ взаимодействует с водой с образованием угольной кислоты: СО2 + Н2О За счет угольной кислоты возникает избыток ионов водорода, который связывается с калиевой солью гемоглобина, отдавшей кислород: КbHb + Н2СО3 В результате этого процесса накапливаются анионы
Взаимодействие буферных систем и физиологических механизмов в регуляции КЩР. Кислые продукты нейтрализуются бикарбонатной и гемоглобиновой системами. Буферные основания Роль почек в регуляции КЩР. В клетках эпителия почечных канальцев из СО2 и Н2О под действием карбоангидразы образуется угольная кислота, которая диссоциирует на ионы Н+ и Судьба ионов Н+, попавших в мочу. 1. Н+ + 2. Н+ + 3. Н+ + Ионы
По механизму 1. Метаболический 2. Респираторный (газовый) Метаболический ацидоз – много кислых продуктов (кетоновые тела, лактат, пируват). Наблюдается при голодании, диабете, лихорадке, нарушении кровообращения, поражении почек. Респираторный (газовый) ацидоз наблюдается при заболеваниях легких (бронхиты, пневмонии), недостаток кровообращения. По компенсации: 1. Компенсаторный, рН в N, но меняются другие показатели КЩР. 2. Декомпенсированный – ацидотическая кома. Алкалоз – состояние дефицита ионов водорода в крови. Метаболический алкалоз – дефицит ионов Н+ в крови, сочетается со снижением содержания ионов К+ в крови. Компенсация метаболического алкалоза осуществляется дыхательным и почечным путем (снижение вентиляции с повышением СО2 и увеличением экскреции бикарбоната почками). Причины: дефицит калия, понос, рвота, инфекции ЖКТ, длительное введение стероидных гормонов. Бывает компенсированный и реже декомпенсированный. Дыхательный алкалоз – избыточное выведение Н2СО3. возникает при быстром удалении СО2 и сопровождается развитием гипокапнии (понижение парциального давления СО2 в артериальной крови – менее 35 мм рт. столба).дыхательный алкалоз наблюдается при вдыхании чистого кислорода, при подъеме на высоту, при компенсаторной одышке. Бывает компенсированный.
Причины гипопротеинемий Задержка воды в организме при отеках на почве цирроза печени, потеря белков с мочой при нефритах, угнетение синтеза белков (при длительных инфекционных заболеваниях, при раковой кахексии, при голодании). Причины гиперпротеинемий Потеря больших количеств воды при диарее, неукротимой рвоте, тяжелых травмах ожогах. Это относительная гиперпротеинемия. Абсолютная гиперпротеинемия обусловлена резким нарастанием иммуноглобулинов и патологических белков. (миеломная болезнь).
Строение молекулы Ig. Молекула Ig состоит из 4 полипептидных цепей: 2 тяжелых – Н-цепи и 2 легких – L-цепи. Легкие и тяжелые цепи соединены дисульфидными мостиками. Легкие цепи Ig бывают 2 видов: каппа и лямбда. Тяжелые цепи определяют специфичность Ig и различаются по аминокислотному составу. Различают 5 типов тяжелых цепей: IgG – Для связывания АГ на молекуле Ig имеются активные центры. А.ц. образован V-участками легкой и тяжелой цепи. Разрыв S-S между цепями приводит к исчезновению активных центров. На молекуле Ig два активных центра, т.е. Ig бивалентны. А.ц. – щель, которая комплементарна молекуле АГ. Комплементарность стереоспецифическая и электрохимическая. В молекуле Ig имеется шарнирный участок – это середина Н-цепей. Шарнирный участок не имеет вторичной структуры, здесь находится пролин и молекула Ig делает изгиб, что обеспечивает приспособление Ig к взаимодействию с АГ. Вторичная структура Ig представлена Ig расщепляются протеолитическими ферментами: папаином и пепсином. Папаин расщепляет молекулу Ig в шарнирной области на 3 фрагмента: 2 Fab и Fc-фрагмент. Fab-фрагмент имеет активный центр и это обеспечивает его связывание с АГ, происходит образование комплекса АГ-АТ. Fc-фрагмент выполняет эффекторную функцию: обеспечивает прохождение через плаценту, прохождение через мембрану клеток, обеспечивает фиксацию иммуноглобулинов на поверхности собственных клеток, связывает комплемент, который способен лизировать клетку. IgG составляет 70-75% общего количества иммуноглобулинов. Концентрация IgG в сыворотке достигает нормы к 7-летнему возрасту. Повышенная концентрация IgG наблюдается при инфекциях, при болезнях печени. Пониженная концентрация IgG наблюдается у новорожденных, при недостаточности гуморального иммунитета. IgG – единственный иммуноглобулин, проникающий через плаценту; благодаря этому плод получает материнские антитела. IgM представляет собой пентамер молекулы IgG. Пять молекул, аналогичных IgG, соединены между собой полипептидной цепью J. IgM лучше, чем другие связывает комплемент. IgM синтезируется в ответ на большинство антигенов у детей первого года жизни. Обнаружение у плода антител класса М указывает на внутриматочную инфекцию. IgA сывороточный иммуноглобулин А бывает в разных формах: обычной, димерной и трехмерной. IgA, которые выходят из кровяного русла через слизистые оболочки в просвет дыхательных путей, помимо J-цепи, имеют секреторный компонент – SC. SC-структура защищает IgA от разрушающего действия ферментов, находящихся в скретах слизистых оболочек. Содержание IgA достигает нормы к 10 годам. IgA – типичный иммуноглобулин слизистых оболочек. Функция IgA – защита слизистых оболочек от инфекций. IgE – содержится в крови в низких концентрациях. IgE обладает высокой цитофильностью, т.е. способностью присоединяться к тучным клеткам и базофилам. Присоединение антигена к IgE, находящаяся на этих клетках, приводит к выделению гистамина и это вызывает развитие реакций гиперчувствительности немедленного типа (аллергических реакций). IgD – не связывает комплемент, не проходит через плаценту, не связывается с тканями. Биологическая функция не ясна; вероятно, он служит рецептором В-лимфоцитов. Динамика выработки антител. IgM являются антителами первичного иммунного ответа. Через несколько дней происходит переключение синтеза IgM на IgG. При вторичном ответе сразу синтезируются IgG. Материнские IgG с периодом полураспада 25 дней проникающие в кровь плода через плаценту, обеспечивают пассивный иммунитет новорожденных, к 3 месяцу из содержание уменьшается вдвое. Синтез собственных IgM осуществляется плодом с 5 недели внутриутробного развития. Катаболизм Ig Период полураспада антител зависит от интенсивности обменных процессов и возраста, у новорожденных это 30-35 дней, затем этот срок сокращается. Ig разрушаются под влиянием тканевых катепсинов на Fc и 2 Fab-фрагмента, небольшое количество около 10% IgG выводится в свободном виде, активный центр сохраняется и по мере выведения продукты катаболизма оказывает защитное действие, связывая АГ. Патология обмена Ig 1. Гипогаммаглобулинемия – снижение уровня всех классов Ig. Бывает 3 видов: 1. Транзитная – у детей. 2. Врожденная. 3. Приобретенная (инфекция, интоксикация, нарушение синтеза Ig, ускоренное разрушение, потеря с мочой). 2. Дисгаммаглобулинемия – снижение (изменение) соотношения между классами Ig. Виды: 1. Врожденная. 2. Приобретенная. I тип – нет G, A, > M II тип – нет M, A, >A III тип – N G,M < A
Белки плазмы крови. Кровь состоит из плазмы и взвешенных в ней форменных элементов. Плазма составляет около 55% от объема крови. Эритроциты составляют основную массу форменных элементов – 44%. Из 9-10% сухого остатка плазмы крови на болю белков приходится 6,5-8,5%. Общее содержание белков составляет 60 – 80 г/л. Для разделения белков плазмы крови используют следующие методы:
Разделение белков проводят с использованием солей щелочных и щелочноземельных металлов. Наиболее часто используют сульфат аммония. Различные белки высаливаются из растворов при разных концентрациях растворов сульфата аммония: альбумины выпадают при 100% насыщении, глобулины при 50%. При действии сульфата аммония, ацетона, спирта белки теряют гидратную оболочку, утрачивают устойчивость и выпадают в осадок. В результате высаливания получают 2 фракции:
На долю фибриногена приходится – 2 – 4 г/л. Плазма, лишенная фибриногена, называется сывороткой. В условиях клинических лабораторий для разделения белков сыворотки крови применяют метод электрофореза. Сыворотка – плазма, не содержащая фибриноген и др. факторы свертывания (т.е. жидкость, которая остается после формирования сгустка крови). Высаливание. Для высаливания применяют соли в разных концентрациях. При малой концентрации солей осаждаются наиболее крупные, тяжелые и обладающие наименьшим зарядом частицы. При повышенной концентрации солей выпадают все более мелкие частицы и устойчивые белковые фракции. При использовании 33%-ного раствора (NH4)2SO4 выпадают белки сыворотки крови, имеющие наибольший молекулярный вес – эйглобулины, при 50%-ной концентрации – псевдоглобулины, при 100%-ной – самые «легкие» альбумины. Белки, осажденные сульфатом аммония, почти не денатурируются; после удаления соли из белкового осадка (диализом через целлофановую мембрану) ее растворяют и используют для различных целей. Чтобы лишить высокомолекулярные частицы устойчивости, необходимо удалить их водные оболочки и снять электрический заряд. При электрофорезе в поддерживающих средах имеет значение размер пор геля (молекулярно-ситовой эффект геля) и размеры молекул белка. Электрофорез на бумаге. Сыворотку крови наносят на бумажную полоску, смоченную буфером. Концы бумажной полосы опущены в раствор электролита, где находятся электроды источника постоянного тока. При замыкании электрической цепи белковые молекулы движутся со скоростью. Пропорциональной величине заряда. За единицу времени проходят разные расстояния, т.е. оказываются на разных участках бумажной полосы. 2. Электорфорез основан на передвижении заряженной частицы в электрическом поле. Белки сыворотки крови различаются по молекулярной массе, заряду и изоэлектрической точке. На основании этих различий белки разделяются в электрическом поле. В зависимости от условий проведения разделения различают электрофорез на бумаге, в растворе и в геле: крахмала, целлюлозы и полиакриламидном геле. а) Электрофорез на бумаге позволяет получить 5 белковых фракций: 1. Альбумины. 2. б) Электрофорез в крахмальном геле позволяет получить 10 фракций. в) Электрофорез в полиакриламидном геле позволяет получить до 18 фракций. При электрофорезе в полиакриламидном геле используют пары буферных растворов с различными значениями рН и различной степени пористости гель. 3. Изоэлектрофокусировние – проведение электрофореза в поддерживающих средах с градиентом рН. Нахождение белка определяется его изоэлектрической точкой. 4. Иммунофорез представляет собой комбинацию электрического и иммунологического методов. Происходит электрофорез и реакция преципитации в одной среде. Иммуноэлектрофорез – сочетание электрофореза в агаровом геле и иммунодиффузии. После электрофоретического разделения белков в канавку, идущую параллельно пути движения антигенов, вносят перциптирующую иммунную сыворотку. Антигены и антитела диффундируют на месте встречи дугообразные полосы преципитации.
Электрофорез основан на способности белков перемещаться в электрическом поле. Для функционирования белков сыворотки крови чаще всего используют веронал-мединаловый буфер с рН 8,6. При этом значении рН белки заряжаются отрицательно и движутся к аноду. В связи с различием в размерах молекул и в величине заряда скорость движения белков самые малые и имеющие наибольший заряд альбумины, затем Скорость передвижения белка в электрическом поле зависит от его электрофоретической подвижности.
|
|||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-08-16; просмотров: 1842; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.119 (0.01 с.) |



 HНbО2
HНbО2
 Н2СО3
Н2СО3 + HHb
+ HHb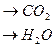
 Н2СО3
Н2СО3 

 - выводится
- выводится
 + Сl + Na
+ Сl + Na  ).
). Нарушения КЩР
Нарушения КЩР


 Нарушения обмена белков плазмы крови
Нарушения обмена белков плазмы крови


 Уменьшение общего содержания белков в сыворотке крови
Уменьшение общего содержания белков в сыворотке крови
 , IgA –
, IgA –  , IgM –
, IgM –  , IgD –
, IgD –  . На легких и тяжелых цепях выделяют вариабельные и константные участки. Вариабельные участки у иммуноглобулинов, направленных против различных АГ различны.
. На легких и тяжелых цепях выделяют вариабельные и константные участки. Вариабельные участки у иммуноглобулинов, направленных против различных АГ различны. -структуры или «клубки». «Клубки» называются доменами, которые возникают при образовании дисульфидных связей между аминокислотными остатками внутри одной цепи.
-структуры или «клубки». «Клубки» называются доменами, которые возникают при образовании дисульфидных связей между аминокислотными остатками внутри одной цепи.




