Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Эмбриональный период, его характеристика. Генный контроль эмбрионального развития.Содержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
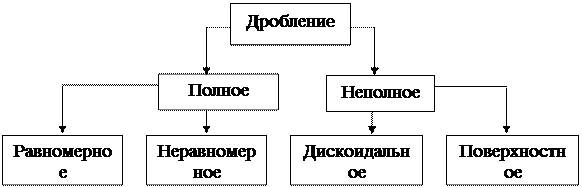
Эмбриональный период (эмбриогенез) начинается с момента проникновения сперматозоида в яйцеклетку, т.е. образования зиготы и заканчивается выходом нового организма из яйцевых оболочек или моментом рождения. В эмбриогенезе различают периоды: предзиготный, зиготный, дробление, гаструляцию, гисто- и органогенез. Предзиготный период характеризуется процессами: - гаметогенез – образование гамет; - амплификация генов – синтез и запасание р-РНК и и-РНК в яйце; - ооплазматическая сегрегация – дифференцировка кортекса цитоплазмы на зоны; - образование кортикального слоя цитоплазмы, содержащего гранулы гликогена; - яйцо приобретает полярность формируются: вегетативный, отягощенный желтком, и анимальный полюса. Процесс оплодотворения у многоклеточных организмов состоит в объединении спермия и яйцеклетки и образовании качественно новой клетки – зиготы. Зигота – это одноклеточный зародыш. На этой стадии геном не активен. Следующий этап – дробление зиготы (Рис. 8).
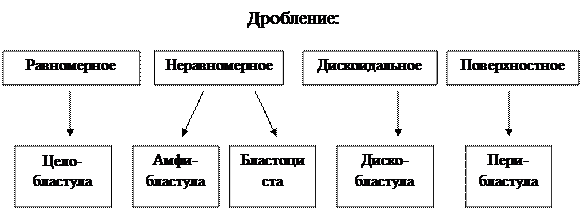
Рис. 8 — Основные типы дробления В основе этого процесса лежит митотическое деление клеток. Однако образующиеся в результате деления дочерние клетки не расходятся, а остаются тесно прилегающими друг к другу. В процессе дробления дочерние клетки прогрессивно уменьшаются. Каждому животному свойствен определенный тип дробления, обусловленный количеством и характером распределения желтка в яйцеклетке. Желток тормозит дробление, поэтому часть зиготы, перегруженная желтком, дробится медленнее или не дробится вовсе. В изолецитальном, бедном желтком оплодотворенном яйце ланцетника, первая борозда дробления в виде щели начинается на анимальном полюсе и постепенно распространяется в продольном меридиональном направлении к вегетативному, разделяя яйцо на 2 клетки – 2 бластомера. Вторая борозда также проходит меридиональном направлении, но перпендикулярно первой – образуются 4 бластомера. Третья борозда проходит экваториально: возникает 8 бластомеров. В результате последующих дроблений в меридиональных и экваториальных плоскостях образуется 16, 32, 64 и т.д. бластомеров. В результате ряда последовательных дроблений формируются группы клеток, тесно прилегающих друг к другу. У некоторых животных такой зародыш напоминает ягоду шелковицы или малины. Он получил название морулы (лат. morum – тутовая ягода) – многоклеточного шара без полости внутри. В телолецитальных яйцах, перегруженных желтком, дробление может быть полным равномерным или неравномерным и неполным. Бластомеры вегетативного полюса из-за обилия инертного желтка всегда отстают в темпе дробления от бластомеров анимального полюса. Полное, но неравномерное дробление характерно для яиц амфибий. У рыб, птиц и некоторых других животных дробится лишь часть яйца, расположенная на анимальном полюсе; происходит неполное дискоидальное дробление. В процессе дробления увеличивается число бластомеров, однако бластомеры не вырастают до размеров исходной клетки, а с каждым дроблением становятся мельче. Это объясняется тем, что митотические циклы дробящейся зиготы не имеют типичной интерфазы; пресинтетический период (G1) отсутствует, а синтетический (S) начинается еще в телофазе предшествующего митоза. Во время дробления митозы следуют быстро друг за другом, и к концу периода весь зародыш лишь не намного крупнее зиготы. В это время бластомеры уже отличаются по характеру цитоплазмы, по содержанию желтка, размерам, что влияет на их дальнейшее развитие и дифференцировку. Дробление яйца заканчивается образованием бластулы. Бластула – это многоклеточный однослойный зародыш. В клетках бластулы устанавливается типичное для каждого вида животных ядерно-плазматическое соотношение. Начиная с бластулы, клетки зародыша называют эмбриональными клетками. У ланцетника бластула образуется по достижении зародышем 128 клеток. В силу накопления продуктов жизнедеятельности бластомеров между ними появляется полость (бластоцель, или первичная полость). При полном равномерном дроблении (как у ланцетника) бластула имеет форму пузырька со стенкой в один слой клеток, который назван бластодермой. Стадию бластулы проходят зародыши всех типов животных. У млекопитающих дробление полное неравномерное, т.к. желтка в яйцах мало. В различных бластомерах оно идет с разным ритмом, и можно наблюдать стадии 2, 3, 6, 7, 9, 10 и т.д. бластомеров. Одни из них (светлые) располагаются по периферии, другие (темные) находятся в центре. Из светлых клеток образуется окружающий зародыш трофобласт, клетки которого выполняют вспомогательную функцию и непосредственно в формировании тела зародыша не участвуют. Клетки трофобласта обладают: 1) способностью растворять ткани, благодаря чему зародыш внедряется в стенку матки; 2) отслаиваются от клеток зародыша, образуя полый пузырек. Полость трофобласта заполняется жидкостью, диффундирующей в нее из тканей матки. Зародыш в это время имеет вид узелка, расположенного на внутренней стенке трофобласта. В результате дальнейшего дробления зародыш принимает форму диска, распластанного на внутренней поверхности трофобласта. Типы бластул зависят от типа дробления (Рис. 9).
Рис. 9 — Зависимость типов бластул от дробления У всех многоклеточных животных следующим за бластулой этапом развития является гаструляция, которая представляет собой процесс перемещения эмбрионального материала с образованием двух или трех слоев тела зародыш, называемых зародышевыми листками. В процессе гаструляции следует различать два этапа: 1) образование экто- и энтодермы – двухслойный зародыш; 2) образование мезодермы – трехслойный зародыш. У животных с изолецитальным типом яиц гаструляция идет путем инвагинации, т.е. впячивания. Вегетативный полюс бластулы впячивается внутрь. Противоположные полюса бластодермы почти смыкаются, так что бластоцель либо исчезает полностью, либо остается в виде незначительной полости, а из шара возникает двухслойный зародыш. Внешний слой клеток носит название эктодермы. Внутренний слой называется энтодерма. Формирующаяся полость называется гастроцелем, или первичной кишкой, а вход в кишку получил наименование бластопора, или первичного рта. Края его сближаются, образуя верхнюю и нижнюю губы. У первичноротых, к которым относится большинство типов беспозвоночных, бластопор превращается в окончательный рот, у вторичноротых (иглокожих и хордовых) из него формируется анальное отверстие либо он зарастает, а рот образуется на противоположном конце тела. Другими способами гаструляции являются: деляминация, эпиболия, иммиграция и смешанный способ. При деляминации клетки зародыша делятся параллельно его поверхности, образуя наружный и внутренний зародышевый листки. Эпиболия встречается у животных, имеющих телолецитальные яйца. При этом способе гаструляции мелкие клетки анимального полюса обрастают и покрывают снаружи крупные, богатые желтком клетки вегетативного полюса, которые становятся внутренним слоем. Образование гаструлы путем иммиграции характерно для кишечнополостных животных. Этот способ заключается в массовом активном перемещении клеток бластодермы в бластоцель. Чаще всего имеет место смешанный тип гаструляции, когда одновременно проходят и впячивание, и обрастание, и миграция. Так протекает, например, гаструляция у земноводных. На стадии двух зародышевых листков заканчивается развитие губок и кишечнополостных. У всех организмов, относящихся к типам, стоящим на более высоких ступенях эволюции, развиваются три зародышевых листка. Третий, или средний зародышевый листок называется мезодермой, так как он образуется между наружным и внутренним листками. Различают два основных способа образования мезодермы – телобластический и энтероцельный. Телобластический способ встречается у многих беспозвоночных. Заключается он в том, что вблизи бластопора с двух сторон первичной кишки во время гаструляции образуется по одной крупной клетке телобласту. В результате размножения телобластов формируется мезодерма. Энтероцельный способ характерен для хордовых. В этом случае с двух сторон от первичной кишки образуются выпячивания – карманы, или целомические мешки. Внутри карманов находится полость, представляющая собой продолжение первичной кишки – гастроцеля. Целомические мешки полностью отшнуровываются от первичной кишки и разрастаются между экто- и энтодермой. Клеточный материал этих участков дает начало мезодерме. Дорсальный отдел мезодермы, лежащий по бокам от нервной трубки и хорды, расчленен на сегменты – сомиты. Вентральный ее отдел образует сплошную боковую пластинку, находящуюся по бокам кишечной трубки. Сомиты дифференцируются на три отдела: медиальный – склеротом, центральный – миотом и латеральный – дерматом. В вентральной части мезодермальной закладки принято различать нефрогонотом (ножка сомита) и спланхнотом. Закладка спланхнотома разделяется на два листка, между которыми образуется полость. Она называется вторичной полостью, или целомом. Висцеральный листок граничит с энтодермальной кишечной трубкой, а париентальный – лежит непосредственно под эктодермой. Гистогенез – процесс образования тканей. Органогенез – формирование органов. Дифференцированный на три эмбриональных листка зародышевый материал дает начало всем тканям и органам. Из эктодермы развиваются ткани нервной системы, которая очень рано обособляется. У хордовых она первоначально имеет форму нервной пластинки. Эта пластинка растет интенсивнее остальных участков эктодермы и затем прогибается, образуя желобок. Размножение клеток продолжается, края желобка смыкаются, возникает нервная трубка, которая тянется вдоль тела от переднего конца к заднему. На переднем конце нервной трубки путем дальнейшего роста и дифференцировки формируется головной мозг. Отростки нервных клеток центральных отделов нервной системы образуют периферические нервы. Кроме того, из эктодермы развиваются наружный покров кожи – эпидермис и его производные (ногти, волосы, сальные и потовые железы, эмаль зубов, воспринимающие клетки органов зрения, слуха, обоняния и т.п.). Из энтодермы развивается эпителиальная ткань, выстилающая органы дыхательной, частично мочеполовой и пищеварительной систем, в том числе печень и поджелудочную железу. Из мезодермы: м иотом дает начало скелетной мускулатуре, нефрогонотом – органам выделения и половым железам (гонадам). Клетки, образующие висцеральные и париетальные листки спланхнотома, являются источником эпителиальной выстилки вторичной полости тела – целома. За счет элементов склеротома развиваются хрящевая, костная и соединительная ткани внутренних органов, кровеносных сосудов, гладкой мускулатуры кишок, дыхательных и мочеполовых путей. В образовании сердца принимает участие также висцеральный листок спланхнотома. Дерматом дает начало соединительной ткани кожи. Железы внутренней секреции имеют различное происхождение: одни из них (эпифиз, часть гипофиза) развиваются из закладок нервной системы, другие – из эктодермы. Надпочечники и половые железы являются производными мезодермы. Органогенез. В этой стадии можно выделить две фазы. Первая – нейруляция, состоит в образовании комплекса осевых органов: нервной трубки, хорды и кишечника. Зародыш на стадии нейруляции называется нейрулой. Нервная трубка образуется в результате опускания пласта клеток эктодермы, образования вначале нервного желобка, края которого затем смыкаются. Передний, расширенный отдел в дальнейшем развитии образует головной мозг, остальная часть нервной трубки спинной мозг. Отличительной чертой первой фазы органогенеза служит то, что в морфологические перестройки, сопровождающиеся формированием центральной нервной системы, вовлекается почти весь зародыш. Вторая фаза заключается в построении остальных органов, приобретении различными участками тела типичной для них формы и черт внутренней организации, установлении определенных пропорций. Развитие других органов представляет собой пространственно ограниченные процессы. Образование хорды по времени соответствует самым ранним этапам нейруляции и происходит путем обособления по средней линии клеточного материала из зачатка, общего с энтодермой и мезодермой – стенки первичной кишки. Органогенез завершается в основном к концу эмбрионального периода развития. Однако дифференцировка и усложнение органов продолжается и в постэмбриональном онтогенезе. Описанные процессы связаны не только с активным клеточным размножением первичных эмбриональных закладок, но и с их значительным перемещением, изменением формы тела зародыша, образованием отверстий и полостей, а также с формированием ряда временных зародышевых (провизорных) органов. Провизорные органы. Назначение провизорных органов – обеспечение жизненных функций зародыша в разнообразных условиях среды. Эмбриональное развитие организмов с разным типом онтогенеза протекает в различных условиях. Во всех случаях развития необходимая связь зародыша со средой обеспечивается специальными внезародышевыми органами, функционирующими временно и называемыми провизорными. Степень развития и функция этих органов различны. Для всех животных с неличиночным типом развития, яйца которых богаты желтком (рыбы, рептилии, птицы), характерен такой провизорный орган, как желточный мешок. В стенки желточного мешка врастают кровеносные сосуды, образующие по всей поверхности желтка густую капиллярную сеть. Клетки стенки желточного мешка выделяют ферменты, расщепляющие питательные вещества желтка, которые поступают в кровеносные капилляры и далее в организм зародыша. Желточный мешок – это также первый кроветворный орган зародыша, место размножения клеток крови. У млекопитающих редуцированный желточный мешок входит в состав пупочного канатика. Амнион развивается у истинно наземных животных. Амнион осуществляет функции обмена и защиты от высыхания и механических воздействий. Амниотическая жидкость, в которую погружен развивающийся эмбрион, представляет собой водный раствор белков, сахаров, минеральных солей, содержит также гормоны и мочевину. В процессе развития состав этой среды изменяется. В акушерской практике амниотическую жидкость, отходящую перед родами, называют водами. Позвоночные, обладающие амнионом (пресмыкающиеся, птицы и млекопитающие), объединяются в группу высших позвоночных, или амниотов. Низшие позвоночные, не имеющие амниона (круглоротые, рыбы и земноводные), составляют группу анамний. Аллантоис – вырост задней кишки зародыша. Наиболее развит у животных развивающихся в яйце – рептилии и птицы, где служит местом накопления азотистых отходов метаболизма. Аллантоис сливается с хорионом и образуется хорион-аллантоисная оболочка, богатая кровеносными сосудами, через которую эмбрион поглощает кислород, отдает углекислоту и продукты обмена. Хорион или ворсинчатая оболочка выполняет функцию наружной зародышевой оболочки. Названа она так вследствие развития на ее поверхности большого числа выростов, ворсинок. Ворсинки хориона врастают в слизистую оболочку матки. Место наибольшего разветвления ворсинок хориона и наиболее тесного контакта их со слизистой оболочкой матки носит название детского места или плаценты. У млекопитающих и человека яйцеклетка бедна желтком, поэтому провизорные приспособления развивающегося организма имеют свои особенности. Желточный мешок закладывается на ранних этапах эмбриогенеза, но не развивается, а постепенно редуцируется, расслаивается. Аллантоис также не развит. Зачаток его входит в состав нового специфического провизорного органа – пупочного канатика. Генный контроль эмбрионального развития. Основу процесса онтогенеза составляет наследственная информация, получаемая от родителей. Ее реализация зависит от условий внешней и внутренней среды. Общая схема онтогенетических процессов включает три этапа: 1. Информация для экспрессии и репрессии генов. Гены получают информацию от соседних клеток, продуктов метаболизма, гормонов и других факторов для своей активации. 2. Информация от генов в ходе процессов транскрипции и трансляции для синтеза полипептидов. 3. Информация от белков для формирования тканей и органов. В течение онтогенеза в будущей яйцеклетке (овоцитах) происходит синтез р-РНК, рибосом, т-РНК, необходимых для начального периода развития (дробление, образование бластулы). Происходит амплификация генов (умножение) р-РНК в цитоплазме, и-РНК. Хромосомы при этом имеют вид «ламповых щеток». На этих петлях происходит синтез и-РНК. Гены р‑РНК используются для синтеза белковых молекул рибосом клетки. Гены и-РНК используются для трансляции в течение длительного времени (запасаются). Гены сперматозоида при этом не функционируют. За счет функции плазмогенов плазмотипа наблюдается дифференцировка кортекса цитоплазмы яйцеклетки на зоны (ооплазматическая сегрегация). При оплодотворении, вносится геном сперматозоида в яйцеклетку. Генотип зиготы не активен за счет полной репрессии генов. Функция плазмогенов плазмотипа вызывает усиление дифференцировки кортекса на зоны. Дробление регулируется на первых порах исключительно информацией, содержащейся в яйце. Запасание в ооцитах и-РНК обеспечивает активный синтез белка. Геном матери и отца в этот период не транскрибируется. На стадии бластулы активизируется геном сперматозоида. Генетическая информация бластомеров обеспечивает синтез белков. Вплоть до поздней бластулы реализуется та часть генетической информации, которая касается общих метаболических процессов для всех делящихся клеток. Затем наблюдается репрессия тканеспецифических генов, т.е. дифференцировка клеток зародыша. Гаструляция контролируется за счет генетической информации эмбриональных клеток. Функционирование определенных генов по принципу экспрессии и репрессии обеспечивает синтез белков и закладку зародышевых листков эмбриона. Гисто- и органогенез обеспечивается за счет генной информации клеток эмбриона. В этот период обосабливаются стволовые клетки. Разные популяции которых дают начало различным органам и тканям. Процесс контролируется функцией определенных генов по принципу экспрессии и репрессии. В ходе развития устанавливаются индукционные отношения между тканями и органами, т.е. влияние одной ткани на другую, направляющее ее развитие. Так, зачаток хорды контактирует с эктодермой, в результате чего эпидермальные клетки дифференцируются не в эпителий кожи, а в нервную трубку. Это явление называется эмбриональной индукцией. Предполагается, продукты деятельности генов клеток, зачатки хорды активируют деятельность участков хромосом клеток энтодермы, которые определяют развитие нервной системы. Формированием фенотипа зависит от реализации наследственной программы в конкретных условиях среды.
|
|||||||||||
|
|
Последнее изменение этой страницы: 2016-08-01; просмотров: 555; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.226.96.142 (0.012 с.) |