Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Существует два типа нуклеиновых кислот: днк и рнк.Содержание книги
Поиск на нашем сайте
РНК (рибонуклеиновая кислота), так же как ДНК, представляет собой полимер мономерами которого служат нуклеотиды. Азотистые основания те же самые, что входят в состав ДНК (аденин, гуанин, цетозин); четвертое - урацил - присутствует в молекуле РНК вместо тимина. Нуклеотиды РНК содержат вместо дизоксирибозы другую пентозу - рибозу. ДНК (дезоксирибонуклеиновая кислота) - биологический полимер, состоящий из двух полинуклеотидных цепей, соединенных друг с другом. Мономеры, составляющие каждую из цепей ДНК, представляют собой сложные органические соединения, включающие одно из четырех азотистых оснований: аденин (А) или тимин (Т), цитозин (Ц) или гуанин (Г); пятиатомный сахар пентозу - дезоксирибозу, по имени которой получила название и сама ДНК, а также остаток фосфорной кислоты. Эти соединения носят название нуклеотидов. В каждой цепи нуклеотиды соединяются путем образования ковалентных связей между дезоксирибозой одного и остатком фосфорной кислоты последующего нуклеотида. Объединяются две цепи в одну молекулу при помощи водородных связей, возникающих между азотистыми основаниями, входящими в состав нуклеотидов, образующих разные цепи. Исследуя нуклеотидный состав ДНК различного происхождения, Чаргафф обнаружил следующие закономерности. 1. Все ДНК независимо от их происхождения содержат одинаковое число пуриновых и пиримидиновых оснований. Следовательно, в любой ДНК на каждый пуриновый нуклеотид приходится один пиримидиновый. 2. Любая ДНК всегда содержит в равных количествах попарно аденин и тимин, гуанин и цитозин, что обычно обозначают как А =Т и G=C. Из этих закономерностей вытекает третья. 3. Количество оснований, содержащих аминогруппы в положении 4 пиримидинового ядра и 6 пуринового (цитозин и аденин), равно количеству оснований, содержащих оксо-группу в тех же положениях (гуанин и тимин), т. е. A+C=G+T. Эти закономерности получили название правил Чаргаффа. Наряду с этим было установлено, что для каждого типа ДНК суммарное содержание гуанина и цитозина не равно суммарному содержанию аденина и тимина, т. е. что (G+C)/(A+T), как правило, отличается от единицы (может быть как больше, так и меньше ее). По этому признаку различают два основных типа ДНК: А-Т-тип с преимущественным содержанием аденина и тимина и G-C-тип с преимущественным содержанием гуанина и цитозина. Величину отношения содержания суммы гуанина и цитозина к сумме содержания аденина и тимина, характеризующую нуклеотидный состав данного вида ДНК, принято называть коэффициентом специфичности. Каждая ДНК имеет характерный коэффициент специфичности, который может изменяться в пределах от 0,3 до 2,8. При подсчете коэффициента специфичности учитывается содержание минорных оснований, а также замены основных оснований их производными. Например, при подсчете коэффициента специфичности для ЭДНК зародышей пшеницы, в которой содержится 6% 5-метилцитозина, последний входит в сумму содержания гуанина (22,7%) и цитозина (16,8%). Смысл правил Чаргаффа для ДНК стал понятным после установления ее пространственной структуры. Гидролиз РНК (мягкая щелочная среда):
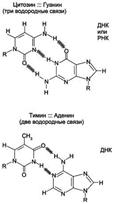
Кислотный гидролиз гетероциклические основания + рибоза ДНК подвергается ферментативному гидролизу под действием нуклеаз. Вторичная структура РНК и ДНК: Вторичная структура ДНК. В 1953 г. Дж. Уотсоном и Ф. Криком была предложена модель пространственной структуры ДНК. Согласно этой модели, молекула ДНК имеет форму спирали, образованную двумя полинуклеотидными цепями, закрученными относительно друг друга и вокруг общей оси. Двойная спираль правозакрученная, полинуклеотидньхе цепи в ней антипараллельны,т.е. если одна из них ориентирована в направлении 3'→5', то вторая - в направлении 5'→3'. Поэтому на каждом из концов молекулы ДНК расположены 5'-конец одной цепи и 3'-конец другой цепи. Все основания цепей ДНК расположены внутри двойной спирали, а пентозофосфатный остов - снаружи. Полинуклеотидные цепи удерживаются относительно друг друга за счёт водородных связей между комплементарными пуриновыми и пиримидиновыми азотистыми основаниями А и Т (две связи) и между G и С (три связи) (рис. 4-7). При таком сочетании каждая
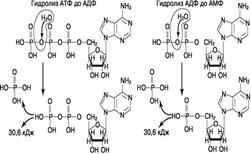
Рис. 4-7. Пурин-пиримидиновые пары оснований в ДНК. пара содержит по три кольца, поэтому общий размер этих пар оснований одинаков по всей длине молекулы. Водородные связи при других сочетаниях оснований в паре возможны, но они значительно слабее. Последовательность нуклеотидов одной цепи полностью комплементарна последовательности нуклеотидов второй цепи. Поэтому, согласно правилу Чаргаффа (Эрвин Чаргафф в 1951 г. установил закономерности в соотношении пуриновых и пиримидиновых оснований в молекуле ДНК), число пуриновых оснований (А + G) равно числу пиримидиновых оснований (Т + С). Комплементарые основания уложены в стопку в сердцевине спирали. Между основаниями двухцепочечной молекулы в стопке возникают гидрофобные взаимодействия, стабилизирующие двойную спираль. Такая структура исключает контакт азотистых остатков с водой, но стопка оснований не может быть абсолютно вертикальной. Пары оснований слегка смещены относительно друг друга. В образованной структуре различают две бороздки - большую, шириной 2,2 нм, и малую, шириной 1,2 нм. Азотистые основания в области большой и малой бороздок взаимодействуют со специфическими белками, участвующими в организации структуры хроматина. 35 Строение АТФ, АДФ, АМФ
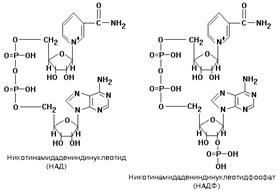
Ферменты, принимающие непосредственное участие в отнятии водорода от субстратов, в настоящее время называются дегидрогеназами Небелковая часть этих ферментов представляет собой динуклеотид: никотинамид-адениндинуклеотид (НАД+) или никотинамидадениндинуклеотидфосфат (НАДФ+). НАД+ и НАДФ+ входят в состав каталитического центра НАДГ. Они являются коферментами, так как связаны с белковой частью слабыми типами связей - могут легко диссоциировать. Они присоединяются к белковой части только в момент протекания реакции. Реакция, которую катализируют НАДГ - это реакция окисления субстрата. Известно около 150 НАДГ, которые различаются по строению белковой части (апофермента). Апоферменты большей части НАДГ способны присоединять или только НАД, или только НАДФ, и лишь немногие способны соединяться и с тем, и с другим коферментами. НАДГ, участвующие в митохондриальном окислении, находятся в матриксе митохондрий, в отличие от большинства других участников дыхательной цепи, которые встроены во внутреннюю мембрану. НАДГ можно встретить и в цитоплазме клеток. Мембрана митохондрий непроницаема для НАД(Ф), поэтому митохондриальный и цитоплазматический НАД(Ф) никогда не смешиваются. В митохондриях содержится очень много НАД и почти нет НАДФ, а в цитоплазме - наоборот - очень много НАДФ и почти нет НАД. Из матрикса митохондриальный НАД×Н2 отдает два атома водорода на «комплекс I», встроенный во внутреннюю мембрану митохондрий. КОМПЛЕКС I В составе комплекса находится 26 полипептидных цепей общей массой 800 кДа. Комплекс содержит следующие небелковые компоненты: Флавинмононуклеотид (ФМН), 5 центров FeS (железо-серные центры): FeS1a, FeS1b FeS2, FeS3, FeS4. В транспорте водорода по дыхательной цепи в этом комплексе принимает участие ФМН. Одновременно с протонами транспортируются и электроны. Наибольшие перепады редокс-потенциала наблюдаются между железо-серными белками, расположенными в следующем порядке: ФМНÞFeS1aÞFeS1bÞFeS3ÞFeS4ÞFeS2 Комплекс I – интегральный белковый комплекс. Используя энергию, выделяющуюся при переносе электронов по дыхательной цепи, он транспортирует 4 протона из матрикса в межмембранное пространство – комплекс I работает как протонный генератор. Точный механизм этого транспорта до сих пор неизвестен. Далее комплекс I восстанавливает промежуточный переносчик KoQ (убихинон). Это жирорастворимое низкомолекулярное вещество, содержащее длинную изопреновую цепь, не имеет белковой части. КоQ принимает водород от комплекса I. Образовавшийся КоQH2 отдает водород на комплекс III. КОМПЛЕКС III. В своем составе содержит цитохромы – сложные белки, содержащие небелковый компонент - простетическую группу, сходню по строению с небелковой частью гемоглобина – гемом. 1) Цитохромы b, имеющие в своем составе два типа простетических групп тетрапиррольной структуры - «гем». Известно два гема цитохромов: be, обладающий низким окислительно-восстановительным потенциалом и bh с высоким окислительно-восстановительным потенциалом. Строение простетической группы цитохромов группы b, похожей на гем белка гемоглобина, представлено на рисунке. Его необходимо выучить. 2)FeSIII – железо-серный кластер. 3) Цитохром С1. Имеет в своем составе особый гем типа «с». Друг от друга цитохромы могут отличаться: 1) Строением белковой части; 2) Значением окислительно-восстановительного потенциала; 3) Строением радикалов, расположенных по периферии гема; 4) Присоединением гема к белковой части – в некоторых случаях гем присоединен к ней ковалентной связью за счет радикалов цистеина, что характерно для цитохромов c1 и c. От двух атомов водорода, которые переносятся на комплекс III от KoQ, дальше по цепи транспортируются только электроны, два протона (H+)комплекс III выбрасывает в межмембранное пространство вместе с еще одной парой протонов, которые подхватываются комплексом из матрикса. Таким образом, комплекс III в сумме выбрасывает в межмембранное пространство 4 протона. Поэтому комплекс III, как и комплекс I, является протонным генератором, и целью его работы также является создание DmH+. КОМПЛЕКС IV. Комплекс IV называется цитохромоксидазой. Он способен захватывать из матрикса 4 протона. Два из них он отправляет в межмембранное пространство, а остальные передает на образование воды.
|
|||||
|
|
Последнее изменение этой страницы: 2016-08-01; просмотров: 588; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.217.242.39 (0.008 с.) |

 РНК щелочн.гидролиз Рибонуклеотиды щелочн.гидролиз Рибонуклеозиды +Н3РО4
РНК щелочн.гидролиз Рибонуклеотиды щелочн.гидролиз Рибонуклеозиды +Н3РО4