Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Транспорт углекислого газа кровью.Содержание книги
Поиск на нашем сайте
Как и кислород, диоксид углерода в крови находится в двух состояниях – физически растворенном и химически связанном. Около 5% СО2 транспортируется в растворенном виде. Химическая связь диоксида углерода осуществляется по реакции СО2 + Н2О↔Н2СО3↔Н+ + НСО3- Реакция сдвигается вправо при высоких напряжениях СО2, и влево при низких. Катализируется карбоангидразой с коэффициентом ускорения 250-300 раз. 80% образовавшейся угольной кислоты транспортируется в виде бикарбонатов щелочных металлов. Противоионами для карбонатных ионов в плазме выступают Na+, а в эритроитах – К+. Остальные 20% НСО3- транспортируется в связи с гемоглобином. В артериальной крови 15% СО2 (в венозной 20%) переносится в виде карбаминовых групп гемоглобина, поскольку NH-группы белка связывают СО2 обратимо. Доля транспорта в связи с гемом ничтожна. Для образования бикарбонатов щелочных металлов используется их резерв, ассоциированный с молекулой гемоглбина. Гемоглобин, как и все белковые молекулы, является амфотерным соединением. В слабощелочной среде (рН 7,35-7,4) гемоглобин и оксигемоглобин ведут себя как слабые кислоты, ассоциируя ионы калия. В артериальной крови 67% НСО3- растворено в плазме, уравновешивающим ионом выступает натрий. Гемоглобин в дезоксиформе слабее по кислотным свойствам, чем оксигемоглобин, поэтому легко отдает К+, при этом реализуется эффект Холдена: оксигенация гемоглобина облегчает отдачу СО2 кровью, а дезоксигенация гемоглобина усиливает поглощение диоксида углерода. Гемоглобиновая буферная система (отвечает за 75% буферных свойств крови) и карбонатная буферная система, кроме дыхательной функции, обеспечивает постоянство активной реакции крови в диапазоне рН 7,35 – 7,47 (венозная кровь в норме имеет более кислую реакцию на 0,02 единицы рН). Поэтому нарушения кислотно-щелочного равновесия в организме может быть не только метаболическим, но и дыхательным. При респираторном ацидозе рН крови снижается, концентрация НСО3- возрастает. При респираторном алкалозе (может наступить при гипервентиляции) наблюдаются обратные процессы, концентрация НСО3-падает из-за «вымывания» углекислого газа. Общее содержание СО2 в 100 мл венозной крови составляет около 52 мл, в артериальной крови и легочных капиллярах только 47-48. Артерио-венозная разница по СО2 составлет 4-5 мл на 100 мл крови.
Регуляция дыхания. Конечная цель регуляции дыхания, или полезный приспособительный результат – поддержание постоянного газового состава и рН артериальной и венозной крови. Отклонение этих показателей от нормы (РО2 менее 100 мс рт.ст., РСО2 более 40 мм рт.ст., рН от 7,36) воспринимается как стимул для регуляции. Координированные сокращения дыхательных мышц обеспечиваются ритмической активностью нейронов дыхательного центра, или, как это принято формулировать по современным представлениям, центрального дыхательного механизма. К дыхательным нейронам относят те нервные клетки, импульсная активность которых меняется в соответствии с фазами дыхательного цикла. Различают инспираторные нейроны (нейроны вдоха) и экспираторные (нейроны выдоха) и клеточные популяции, согласовывающие смену дыхательных фаз. Центральный дыхательный механизм локализован в ретикулярной формации продолговатого мозга. Большинство нейронов сгруппированы в двух главных группах ядер – дорсальной и вентральной. В дорсальной группе сосредоточены инспираторные нейроны, посылающие аксоны в шейные сегменты спинного мозга, где они синаптически оканчиваются на мотонейронах ядра диафрагмального нерва. Ядра вентральной группы дыхательных ядер содержат как инспираторные, так и экспираторные нейроны. Они связаны синаптически с теми нейронами спинного мозга, которые иннервируют межреберные мышцы. Для 80% нейронов грудного отдела спинного мозга характерна дыхательная ритмика. В области моста выделен пневмотаксический центр, клетки которого принимают участие в переключении фаз дыхательного цикла. Для нейронов центрального дыхательного механизма характерен автоматизм, хотя пейсмекеров пока не обнаружено. Основной активатор дыхательного механизма – афферентная сигнализация от рецепторов, расположенных во внутренней среде организма. Главный дыхательный стимул – снижение в крови содержания кислорода и повышение напряжения диоксида углерода. Хеморецепторы посылают в ЦНС сигналы о степени отклонения этих показателей от нормы. Основное место локализации хеморецепторов дыхательной системы – область каротидного синуса (каротидные клубочки). В области дуги аорты расположена вторая группа хеморецепторов, контролирующая газовые и кислотные показатели той порции крови, которая направляется к внутренним органам. В продолговатом мозге имеются и центральные хеморецепторы.
Установлено, что чем выше в крови рСО2, тем выше частота импульсации в афферентных волокнах синокаротидного нерва. Эта афферентная посылка интегрируется центральным дыхательным механизмом и используется для усиления дыхания, как увеличением частоты дыхательных циклов, так и углублением каждого вдоха. В трахее, бронхах имеются собственные рецепторы, инициирующие защитные рефлексы дыхания, например, кашель. Кроме того, часть из них используется и для коррекции частоты и глубины дыхания. К ним относится рефлекс Геринга–Брейера. Рецепторы, реагирующие на повышение давления в воздухоносных путях, активируются при вдохе и посылают афферентные сигналы по волокнам блуждающего нерва к группе нейронов дорсальной порции дыхательного центра. Их возбуждение нарастает в фазу вдоха и тормозит инспираторные нейроны. Каждый вдох за счет рецепторов растяжения подготавливает свое окончание. Имеются рецепторы и в верхних дыхательных путях, они активируются при попадании в нос и рот пыли или ирритантов. Кашель, чихание, принюхивание, остановка дыхания на вдохе при обнаружении резкого неприятного запаха или химическом загрязнении среды – рефлекторные проявления их активации.
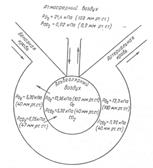
49. Связывание и транспорт газов кровью. Регуляция актов дыхания. Особенности дыхания птиц и рыб. Мы рассмотрели только одну сторону дыхательного процесса — внешнее дыхание, т. е. обмен газов между организмом и окружающей его средой. Местом же потребления кислорода и образования углекислого газа являются все клетки организма, где осуществляется тканевое или внутреннее дыхание. Вследствие этого, когда речь идет о дыхании в целом, необходимо учитывать пути и условия переноса газов: кислорода — от легких к тканям, углекислого газа — от тканей к легким. Посредником между клетками и внешней средой является кровь. Она доставляет тканям кислород и уносит углекислый газ. Движение газов из окружающей среды в жидкость и из жидкости в окружающую среду осуществляется благодаря разности их парциального давления. Газ всегда диффундирует из среды, где имеется высокое давление, в среду с меньшим давлением. Это происходит до тех пор, пока не установится динамическое равновесие. Проследим путь кислорода из окружающей среды в альвеолярный воздух, затем в капилляры малого и большого круга кровообращения и к клеткам организма. Парциальное давление кислорода в атмосферном воздухе 21,1 кПа (158 мм рт. ст.), в альвеолярном воздухе — 14,4—14,7 кПа (108—110 мм рт. ст.) и в венозной крови, притекающей к легким,—5,33 кПа (40 мм рт. ст.). В артериальной крови капилляров большого круга кровообращения напряжение кислорода составляет 13,6—13,9 кПа (102—104 мм рт. ст.), в межтканевой жидкости — 5,33 кПа (40 мм рт. ст.), в тканях — 2,67 кПа (20 мм рт. ст.) и меньше в зависимости от функциональной активности клеток. Таким образом, на всех этапах движения кислорода имеется разность его парциального давления, что способствует диффузии газа. Движение углекислого газа происходит в противоположном направлении. Напряжение углекислого газа в тканях, в местах его образования — 8,0 кПа и более (60 и более мм рт. ст.), в венозной крови — 6,13 кПа (46 мм рт. ст.), в альвеолярном воздухе — 0,04 кПа (0,3 мм рт. ст.). Следовательно, разность напряжения углекислого газа по пути его следования является причиной диффузии газа от тканей в окружающую среду. Схема диффузии газов через стенку альвеол представлена на рис. 3. Однако одними физическими закономерностями объяснить движение газов нельзя. В живом организме равенства парциального давления кислорода и углекислого газа на этапах их движения никогда не наступает. В легких постоянно происходит обмен газов вследствие дыхательных движений грудной клетки, в тканях же разность напряжения газов поддерживается непрерывным процессом окисления.
Рис. 3. Схема диффузии газов через мембрану альвеолы Транспорт кислорода кровью. Кислород в крови находится в двух состояниях: физическом растворении и в химической связи с гемоглобином. Из 19 об% кислорода, извлекаемого из артериальной крови, только 0,3 об% находится в растворенном состоянии в плазме, остальная же часть кислорода химически связана с гемоглобином эритроцитов. Гемоглобин образует с кислородом очень непрочное, легко диссоциирующее соединение — оксигемоглобин: 1 г гемоглобина связывает 1,34 мл кислорода. Содержание гемоглобина в крови составляет в среднем 140 г/л (14 г%). 100 мл крови может связать 14х1,34 = 18,76 мл кислорода (или 19 об%), что составляет в основном так называемую кислородную ёмкость крови. Следовательно, кислородная емкость крови представляет собой максимальное количество кислорода, которое может быть связано 100 мл крови. Насыщение гемоглобина кислородом колеблется от 96 до 98%. Степень насыщения гемоглобина кислородом и диссоциация оксигемоглобина (образование восстановленного гемоглобина) не находятся в прямой пропорциональной зависимости от напряжения кислорода. Эти два процесса не являются линейными, а совершаются по кривой, которая получила название кривой связывания или диссоциации оксигемоглобина. При нулевом напряжении кислорода оксигемоглобина в крови нет. При низких значениях парциального давления кислорода скорость образования оксигемоглобина невелика. Максимальное количество гемоглобина (45—80%) связывается с кислородом при его напряжении 3,47—6,13 кПа (26—46 мм рт. ст.). Дальнейшее повышение напряжения кислорода приводит к снижению скорости образования оксигемоглобина. Сродство гемоглобина к кислороду значительно понижается при сдвиге реакции крови в кислую сторону, что наблюдается в тканях и клетках организма вследствие образования углекислого газа. Это свойство гемоглобина имеет важное значение для организма. В капиллярах тканей, где концентрация углекислого газа в крови увеличена, способность гемоглобина удерживать Кислород уменьшается, что облегчает его отдачу клеткам. В альвеолах легких, где часть углекислого газа переходит в альвеолярный воздух, способность гемоглобина связывать кислород вновь возрастает.
Переход гемоглобина в оксигемоглобин и из него в восстановленный зависит и от температуры. При одном и том же парциальном давлении кислорода в окружающей среде при температуре 37—38° С в восстановленную форму переходит наибольшее количество оксигемоглобина. Таким образом, транспорт кислорода обеспечивается, в основном, за счет химической связи его с гемоглобином эритроцитов. Насыщение гемоглобина кислородом зависит в первую очередь от парциального давления газа в атмосферном и альвеолярном воздухе. Одной из основных причин, способствующих отдаче кислорода гемоглобином, является сдвиг активной реакции среды в тканях в кислую сторону. Транспорт углекислого газа кровью. Растворимость углекислого газа в крови выше, чем растворимость кислорода. Однако только 2,5—3 об% углекислого газа из общего его количества (55—58 об%) находится в растворенном состоянии. Большая часть углекислого газа содержится в крови и в эритроцитах в виде солей угольной кислоты (48—51 об%), около 4—5 об% — в соединении с гемоглобином в виде карбгемоглобина, около 2/з всех соединений углекислого газа находится в плазме и около 1/з в эритроцитах. Угольная кислота образуется в эритроцитах из углекислого газа и воды. И.М. Сеченов впервые высказал мысль о том, что в эритроцитах должен содержаться какой-то фактор типа катализатора, который ускоряет процесс синтеза угольной кислоты. Однако лишь в 1935 г. предположение, высказанное И.М. Сеченовым, было подтверждено. В настоящее время установлено, что в эритроцитах содержится угольная ангидраза (карбоангидраза) — биологический катализатор, фермент, который значительно (в 300 раз) ускоряет расщепление угольной кислоты в капиллярах легких. В тканевых же капиллярах при участии карбоангидразы происходит синтез угольной кислоты в эритроцитах. Активность карбоангидразы в эритроцитах настолько велика, что синтез угольной кислоты ускоряется в десятки тысяч раз. Угольная кислота отнимает основания от восстановленного гемоглобина, в результате чего образуются соли угольной кислоты — бикарбонаты натрия в плазме и бикарбонаты калия в эритроцитах. Кроме того, гемоглобин образует химическое соединение с углекислым газом — карбгемоглобин. Впервые это соединение обнаружено И.М. Сеченовым. Роль карбгемоглобина в транспорте углекислого газа достаточно велика. Около 25—30% углекислого газа, поглощаемого кровью в капиллярах большого круга кровообращения, транспортируется в виде карбгемоглобина. В легких гемоглобин присоединяет кислород и переходит в оксигемоголбин. Гемоглобин вступает в реакцию с бикарбонатами и вытесняет из них угольную кислоту. Свободная угольная кислота расщепляется карбоангидразой на углекислый газ и воду. Углекислый газ диффундирует через мембрану легочных капилляров и переходит в альвеолярный воздух. Уменьшение напряжения углекислого газа в капиллярах легких способствует расщеплению карбгемоглобина с освобождением углекислого газа.
Таким образом, углекислый газ переносится к легким в форме бикарбонатов и в состоянии химической связи с гемоглобином (карбгемоглобин). Важная роль в сложнейших механизмах транспорта углекислого газа принадлежит карбоангидразе эритроцитов. Конечной целью дыхания является снабжение всех клеток кислородом и удаление из организма углекислого газа. Для осуществления этой цели дыхания необходим ряд условий: 1) нормальная деятельность аппарата внешнего дыхания и достаточная вентиляция легких; 2) нормальный транспорт газов кровью; 3) обеспечение системой кровообращения достаточного кровотока; 4) способность тканей «забирать» из протекающей крови кислород, утилизировать его и отдавать в кровь углекислый газ. Таким образом, тканевое дыхание обеспечивается функциональными взаимосвязями между системами дыхания, крови и кровообращения. Регуляция дыхания осуществляется путем рефлекторных реакций, возникающих в результате возбуждения специфических рецепторов, заложенных в легочной ткани, сосудистых рефлексогенных зонах и других участках. Центральный аппарат регуляции дыхания представляют образования спинного мозга, продолговатого мозга и вышележащих отделов нервной системы. Основная функция управления дыханием осуществлянется дыхательными нейронами ствола головного мозга, которые передают ритмические сигналы в спинной мозг к мотонейронам дыхательных мышц. Дыхательный нервный центр – это совокупность нейронов центральной нервной системы, обеспечивающих координированную ритмическую деятельность дыхательных мышц и постоянное приспособление внешнего дыхания к изменяющимся условиям внутри организма и в окружающей среде. Основная (рабочая) часть дыхательного нервного центра расположена в продолговатом мозгу. В ней различают два отдела: инспираторный (центр вдоха) и экспираторный (центр выдоха). Дорсальная группа дыхательных нейронов продолговатого мозга состоит преимущественно из инспираторных нейронов. Они частично дают поток нисходящих путей, вступающих в контакт с мотонейронами диафрагмального нерва. Вентральная группа дыхательных нейронов посылает преимущественно нисходящие волокна к мотонейронам межреберных мышц. В передней части варолиева моста обнаружена область, названная пневмотаксическим центром. Этот центр имеет отношение к работе как экспи-, так и инспираторного его отделов. Важной частью дыхательного нервного центра является группа нейронов шейного отдела спинного мозхга (III-IV шейные сегменты), где расположены ядра диафрагмальных нервов. К моменту рождения ребенка дыхательный центр способен давать ритмическую смену фаз дыхательного цикла, но эта реакция очень несовершенна. Дело заключается в том, что к рождению дыхательный центр еще не сформирован, его формирование заканчивается к 5-6 годам жизни. Это подтверждается тем, что именно к этому периоду жизни детей дыхание у них становится ритмичным и равномерным. У новорожденных же оно неустойчиво как по частоте, так и глубине и ритму. У них дыхание диафрагмальное и практически мало отличается во время сна и бодроствования (частота от 30 до 100 в минуту). У детей 1 года количество дыхательных движений днем в пределах 50-60, а ночью – 35-40 в минуту, неустойчивое и диафрагмальное. В возрасте 2-4 лет – частота становится в пределах 25-35 и носит преимущественно диафрагмальный тип. У 4-6 – летних детей частота дыхания 20-25, смешанное – грудное и диафрагмальное. К 7 –14 годам достигает уровня 19-20 в минуту, оно является в это время смешанным. Таким образом, окончательное формирование нервного центра практически относится к этому возрастному периоду. Как же происходит возбуждение дыхательного центра? Один из важнейших путей его возбуждения - это автоматия. Единой точки зрения на природу автоматии нет, но имеются данные о том, что в нервных клетках дыхательного центра возможно возникновение вторичной деполяризации (по принципу диастолической деполяризации в сердечной мышце), которая, достигая критического уровня, и дает новый импульс. Однако одним из основных путей возбуждения дыхательного нервного центра является его раздражение углекислотой. На прошлой лекции мы отметили, что углекислоты много остается в крови, оттекающей от легких. Она и выполняет функцию основого раздражителя нервных клеток продолговатого мозга. Это опосредуется через специальные образования - хеморецепторы, расположенные непосредственно в структурах продолговатого мозга («центральные хеморецепторы»). Они очень чувствительны к напряжению углекислого газа и кислотно-щелочному состоянию омывающей их межклеточной мозговой жидкости. Углекислота может легко диффундировать из кровеносных сосудов головного мозга в спинномозговую жидкость и стимулировать хеморецепторы продолговатого мозга. Это еще один путь возбуждения дыхательного центра. Наконец, его возбуждение может осуществляться и рефлекторно. Все рефлексы, обеспечивающие регуляцию дыхания мы условно подразделяем на: собственные и сопряженные. Собственные рефлексы дыхательной системы – это такие рефлексы, которые берут начало в органах дыхательной системы и в ней же заканчиваются. В первую очередь к этой группе рефлексов следует отнести рефлекторный акт с механорецепторов легких. В зависимости от, локализации и вида, воспринимаемых раздражений, характера рефлекторных ответов на раздражение различают три вида таких рецепторов: рецепторы рпастяжения, ирритантные рецепторы и юкстакапиллярные рецепторы легких. Рецепторы растяжения легких находятся, преимущественно в гладких мышцах воздухоносных путей (трахее, бронхах). Таких рецепторов в каждом легком около 1000 и связаны они с дыхательным центром крупными миелинизированными афферентными волокнами блуждающего нерва с высокой скоростью проведения. Непосредственным раздражителем этого типа механорецепторов является внутреннее напряжение в тканях стенок воздухоносных путей. При растяжении легких во время вдоха частота этих импульсов возрастает. Раздувание легких вызывает рефлекторное торможение вдоха и переход к выдоху. При перерезке блуждающих нервов эти реакции прекращаются, и дыхание становится замедленным и глубоким. Указанные реакции называют рефлексом Геринга-Брейера. Этот рефлекс воспроизводится у взрослого человека, когда дыхательный объем превосходит 1 л (при физической нагрузке, например). Он имеет большое значение у новорожденных. Ирритантные рецепторы или быстро адаптирующиеся механорецепторы воздухоносных путей, рецепторы слизистой оболочки трахеи и бронхов. Они реагируют на резкие изменения объема легких, а также при действии на слизистую трахеи и бронхов механических или химических раздражителей (пылевых частиц, слизи, паров едких веществ, табачного дыма и т.п.). В отличие от легочных рецепторов растяжения ирритантные рецепторы обладают быстрой адаптацией. При попадании в дыхательные пути мельчайших инородных тел (пыли, частиц дыма), активация ирритантных рецепторов вызывает у человека кашлевой рефлекс. Его рефлекторная дуга такова – от рецепторов информация через верхнегортанный, языкоглоточный, тройничный нерв идет к соотвествующим структурам мозга, отвечающим за выдох (срочный выдох – кашель). Если изолированно возбуждаются рецепторы носовых дыхательных путей, то это вызывает другой срочный выдох - чихание. Юкстакапиллярные рецепторы – расположены вблизи капилляров альвеол и дыхательных бронхов. Раздражителем этих рецепторов является повышение давления в малом круге кровообращения, а также увеличение объема интерстициальной жидкости в легких. Это наблюдается при застое крови в малом круге кровообращения, отеке легких, повреждениях легочной ткани (например, при пневмонии). Импульсы от этих рецепторов направляются к дыхательному центру по блуждающему нерву, вызывая появление частого поверхностного дыхания. При заболеваниях вызывает ощущение одышки, затрудненного дыхания. Может быть не только учащенное дыхание (тахипное), но и рефлекторное сужение бронхов. Еще различают большую группу собственных рефлексов, которые берут свое начало от проприорецепторов дыхательной мускулатуры. Рефлекс от проприорецепторов межреберных мышц осуществляется во время вдоха, когда эти мышцы, сокращаясь, посылают информацию через межреберные нервы к экспираторному отделу дыхательного центра и в результате наступает выдох. Рефлекс от проприорецепторов диафрагмы осуществляется в ответ на ее сокращение во времявдоха, в результате информация поступает по диафрагмальным нервам вначале в спинной, а потом в продолговатый мозг в экспираторный отдел дыхательного центра и наступает выдох. Таким образом, все собственные рефлексы дыхательной системы осуществляются во время вдоха и заканчиваются выдохом. Сопряженные рефлексы дыхательной системы – это рефлексы, которые начинаются за ее пределами. К этой группе рефлексов, прежде всего, относится рефлекс на сопряжение деятельности системы кровообращения и дыхания. Такой рефлекторный акт начинается от периферических хеморецепторов сосудистых рефлексогенных зон. Наиболее чувствительные из них находятся в области синокаротидной зоны. Синокаротидный хеморецептивный сопряженный рефлекс – осуществляется при накоплении углекислого газа в крови. Если его напряжение растет, то происходит возбуждение наиболее высоковозбудимых хеморецепторов (а они именно в этой зоне и находятся в синокаротидном тельце), возникающая волна возбуждения идет от них по IX паре черпномозговых нервов и достигает экспираторного отдела дыхательного центра. Возникает выдох, который и усиливает выброс лишней углекислоты в окружающее пространство. Таким образом, система кровообращения (она, кстати, при осуществлении этого рефлекторного акта также работает более интенсивно, возрастает частота сердечных сокращений, скорость кровотока) влияет на деятельность системы дыхания. Другой разновидностью сопряженных рефлексов дыхательной системы является многочисленная группа экстероцептивных рефлексов. Они берут свое начало от тактильных (вспомните реакцию дыхания на осязание, прикосновение), температурных (тепло – увеличивает, холод – уменьшает дыхательную функцию), болевых (слабые и средней силы раздражители – усиливают, сильные – угнетают дыхание) рецепторов. Проприорецептивные сопряженные рефлексы дыхательной системы осуществляются вследствие раздражения рецепторов скелетных мышц, суставов, связок. Это наблюдается при выполнении физической нагрузки. Почему это происходит? Если в состоянии покоя человеку необходимо 200-300 мл кислорода в минуту, то при физической нагрузке этот объем должен значительно возрости. В этих условиях увеличивается и МО, артериовенозная разница по кислороду. Увеличение этих показателей сопровождается повышением потребления кислорода. Далее все зависит от объема работы. Если работа длится 2-3 минуты и мощность ее достаточно велика, то потребление кислорода непрерывно растет с самого начала работы и снижается лишь после ее прекращения. Если же продолжительность работы больше, то потребление кислорода, нарастая в первые минуты, поддерживается в дальнейшем на постоянном уровне. Потребление кислорода возрастает тем более, чем тяжелее физическая работа. Наибольшее количество кислорода, которое организм может поглотить за 1 минуту при предельно тяжелой для него работе, называется максимальное потребление кислорода (МПК). Работа, при которой человек достигает своего уровня МПК, должна длиться не более 3 минут. Существует много способов определения МПК. У не занимающихся спортом или физическими упражнениями людей величина МПК не превышает 2,0-2,5 л/мин. У спортсменов она может быть выше более чем в два раза. МПК является показателем аэробной производительности организма. Эта способность человека совершать очень тяжелую физическую работу, обеспечивая свои энергетические расходы за счет кислорода, поглощаемого непосредственно во время работы. Известно, что даже хорошо тренированный человек может работать при потреблении кислорода на уровне 90-95% от уровня своего МПК не более 10-15 минут. Тот, кто имеет большую аэробную производительность, тот достигает лучших результатов в работе (спорте) при относительно одинаковой технической и тактической подготовленности. Почему при физической работе возникает увеличение потребления кислорода? В этой реакции можно выделить несколько причин: раскрытие дополнительных капилляров и увеличение крови в них, сдвиг кривой диссоциации гемоглобина вправо и вниз, увеличение температуры в мышцах. Для того, чтобы мышцы могли совершать определенную работу, им нужна энергия, запасы которой в них восстанавливаются при доставке кислорода. Таким образом, существует зависимость между мощностью работы и количеством кислорода, которое требуется для работы. То количество крови, которое требуется для работы, называется кислородным запросом. Кислородный запрос может достигать при тяжелой работе до 15-20 л в минуту и более. Однако максимум потребления кислорода в два-три раза меньше. Можно ли выполнить работу, если минутный кислородный запас превышает МПК? Чтобы правильно ответить на этот вопрос, надо вспомнить, для чего используется кислород при мышечной работе. Он необходим для восстановления богатых энергией химических веществ, обеспечивающих мышечное сокращение. Кислород обычно взаимодействует с глюкозой, и она, окисляясь, освобождает энергию. Но глюкоза может расщепляться и без кислорода, т.е. анаэробным путем, при этом тоже выделяется энергия. Кроме глюкозы, есть и другие вещества, способные расщепляться без кислорода. Следовательно, работа мышц может быть обеспечена и при недостаточном поступлении кислорода в организм. Однако в этом солучае образуется много кислых продуктов и для их ликвидации нужен кислород, ибо они разрушаются путем окисления. То количество кислорода, которое требуется для окисления продуктов обмена, образовавшихся при физической работе, называется кислородный долг. Он возникает во время работы и ликвидируется в восстановительном периоде после нее. На его ликвидацию уходит от нескольких минут до полутора часов. Все зависит от длительности и интенсивности работы. Основную роль в образовании кислородного долга составляет молочная кислота. Чтобы продолжить работу при наличии в крови большого ее количества, организм должен иметь мощные буферные системы и его ткани должны быть приспособлены к работе при недостатке кислорода. Такое приспособление тканей служит одним из факторов, обеспечивающих высокую анаэробную производительность. Все это усложняет регуляцию дыхания при физической работе, так как потребление кислорода в организме возрастает и его недостаток в крови приводит к раздражению хеморецепторов. Сигналы от них идут в дыхательный центр, в результате дыхание учащается. При мышечной работе много образуется углекислоты, которая поступает в кровь и она может действовать на дыхательный центр непосредственно черех центральные хеморецепторы. Если недостаток кислорода в крови приводит преимущественно к учащению дыхания, то избыток углекислоты вызывает его углубление. При физической работе оба эти фактора действуют олновременно, вследствие чего происходит и учащение, и углубление дыхания. Наконец, импульсы идущие от работающих мышц, достигают дыхательного центра и усиливают его работу. При функционировании дыхательного центра все отделы его функционально взаимосвязаны. Это достигается следующим механизмом. При накоплении углекислоты возбуждается инспираторный отдел дыхательного центра, от него информация идет в пневматоксический отдел центра, потом к экспираторному его отделу. Последний, кроме того, возбуждается за счет целой гаммы рефлекторных актов (с рецепторов легких, диафрагмы, межреберных мышц, дыхательных путей, хеморецепторов сосудов). Вследствие его возбуждения через специальный тормозный ретикулярный нейрон угнетается деятельность центра вдоха и на смену ему приходит выдох. Так как центр вдоха тормозится, то он не посылает далее импульсы в пневматоксический отдел, а от него прекращается поток информации в центр выдоха. К этому моменту накапливается в крови углекислота и снимаются тормозные влияния со стороны экспираторного отдела дыхательного центра. Вследствие такого перераспределения потока информации возбуждается центр вдоха и наступает на смену выдоху вдох. И все вновь повторяется. Важным элементом в регуляции дыхания является блуждающий нерв. Именно через его волокна идут основные влияния на центр выдоха. Поэтому в случае его повреждения (также как и при повреждении пневматоксического отдела дыхательного центра) дыхание изменяется так, что вдох остается нормальным, а выдох резко затягивается. Такой тип дыхания называют вагус-диспноэ. Мы уже отмечали выше, что при подъме на высоту происходит увеличение легочной вентиляции, обусловленное стимуляцией хеморецепторов сосудистых зон. Одновременно с этим возрастает частота сердечных сокращений и МО. Эти реакции несколько улучшают кислородный транспорт в организме, но не надолго. Поэтому при длительном пребывании в горах по мере адаптации к хронической гипоксии начальные (срочные) реакции дыхания постепенно уступают место более экономному приспособлению газотранспортной системы организма. Так, у постоянных жителей больших высот реакция дыхания на гипоксию оказывается резко ослабленной (гипоксическая глухота) и легочная вентиляция поддерживается почти на том же уровне, что и у живущих на равнине. Зато при длительном проживании в условиях высокогорья возрастает ЖЕЛ, повышается КЕК, в мышцах становится больше миоглобина, в митохондриях усиливается активность ферментов, обеспечивающих биологическое окисление и гликолиз. У людей, живущих в горах, кроме того, понижена чувствительность тканей организма, в частности, центральной нервной системы, к недостаточному снабжению кислородом. На высотах боле 12000 м давление воздуха очень мало и в этих условиях даже дыхание чистым кислородом не решает проблемы. Поэтому при полетах на этой высоте необходимы герметические кабины (самолеты, космические корабли). Человеку иногда приходиться работать и в условиях повышенного давления (водолазные работы). На глубине в крови начинает растворяться азот и при быстром подъеме из глубины он не успевает выделяться из крови, газовые пузырьки вызывают эмболию сосудов. Состояние, возникающее при этом, называется кесонная болезнь. Она сопровождается болями в суставах, головокружением, одышкой, потерей сознания. Поэтому азот в смесях воздуха заменяют нерастворимыми газами (например, гелием). Человек может произвольно задерживать дыхание не более чем на 1-2 минуты. После предварительной гипервентиляции легких эта задержка дыхания увеличивается до 3-4 минут. Однако затяжное, например, ныряние после гипервентиляции таит в себе серъезную опасность. Быстрое падение оксигенации крови может вызвать внезапную потерю сознания, а в этом состоянии пловец (даже опытный) под влиянием стимула, вызванного ростом парциального напряжения углекислоты в крови, может вдохнуть воду и захлебнуться (утонуть). Итак, в заключение лекции я должен Вам напомнить, что здоровое дыхание это – через нос, как можно реже, с задерджкой во время вдоха и, особенно, после него. Удлиняя вдох, мы стимулируем работу симпатического отдела вегетативной нервной системы, со всеми вытекающими отсюда последствиями. Удлиняя выдох, мы удерживаем больше и дольше в крови углекислоту. А это оказыавает положительное влияние на тонус кровеносных сосудов (снижает его), со всеми вытекающими отсюда последствиями. Благодаря этому кислород может в такой ситуации пройти в самые отдаленные сосуды микроциркуляции, препятствуя нарушению их функции и развитию многочисленных заболеваний. Правильное дыхание – это профилактика и лечение большой группы заболеваний не только дыхательной системы, но и других органов и тканей! Дышите на здоровье! Особенности дыхания у птиц обусловлены их образом жизни — полетом и своеобразием анатомического строения дыхательных путей. Эти особенности свойственны всему классу птиц — как летающих, так и нелетающих. У птиц сильно развита грудная клетка, большая грудная кость, вместо реберных хрящей грудные костные ребра подвижно соединены с позвоночными ребрами. Диафрагма у птиц редуцирована, поэтому не имеет большого значения в дыхании. Наружная поверхность легких вдавлена между ребрами и прочно срастается с ними. Свободная поверхность легких гладкая, покрыта плеврой.
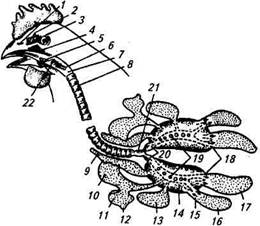
Трахея входит в легкие, разветвляется на бронхи и бронхиолы. В бронхиолах происходит обмен газов с притекающей сюда венозной кровью. Часть бронхов выходит из легких и заканчивается воздухоносными мешками —это тонкостенные образования, заполненные воздухом (рис. 7.11). Они расположены между органами, соединяются с бронхами, а
|
||||||||||||
|
|
Последнее изменение этой страницы: 2021-07-18; просмотров: 146; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.145.37.211 (0.017 с.) |