Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Взаимодействие аллельных геновСодержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
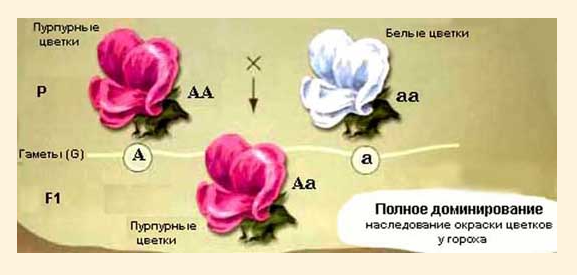
Аллельные гены – различные формы одного и того же гена, расположенные в одинаковых участках (локусах) гомологических хромосом. Аллели определяют варианты развития одного и того же признака. В нормальной диплоидной клетке могут присутствовать не более двух аллелей одного локуса одновременно. Признак, обусловленный взаимодействием одной аллельной пары, называют простым, а форму взаимодействия – аллельной. 1. Полное доминирование (рис. 32) – вид взаимодействия аллельных генов, при котором в гетерозиготном состоянии доминантный аллель полностью подавляет проявление рецессивного аллеля, фенотипически гибриды F1 100% обладают доминантными признаками (гетерозигота имеет фенотип одного из родителей – фенотип доминантной гомозиготы). При наследовании всех менделирующих признаков наблюдается полное доминирование. Менделирующие признаки – признаки, наследующиеся по законам Менделя, например: альбинизм – рецессивный признак, не проявляется в гетерозиготном состоянии. Поэтому у гибридов F2 наблюдается расщепление по генотипу 1:2:1, по фенотипу – 3:1.
Рис. 32. Полное доминирование.
2. Неполное доминирование (рис. 33) – вид взаимодействия аллельных генов, при котором в гетерозиготном состоянии доминантный аллель неполностью подавляет проявление рецессивного аллеля. В результате имеет место промежуточный характер наследования (гетерозигота имеет промежуточный фенотип).
Рис. 33. Неполное доминирование. У гибридов F2 расщепление по генотипу и фенотипу совпадают – 1:2:1.
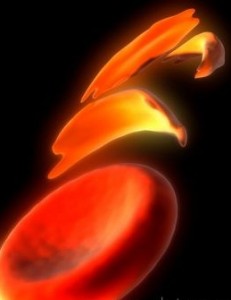
Другим примером неполного доминирования служит серповидноклеточная анемия – наследственная гемоглобинопатия, связанная с таким нарушением строения белка гемоглобина, при котором он приобретает особое кристаллическое строение – так называемый гемоглобин S. Эритроциты, несущие гемоглобин S вместо нормального гемоглобина А, под микроскопом имеют характерную форму серпа (рис. 34), за что эта форма гемоглобинопатии и получила название серповидноклеточной анемии. Эритроциты, несущие гемоглобин S, обладают пониженной стойкостью и пониженной кислород-транспортирующей способностью, поэтому у больных с серповидноклеточной анемией повышено разрушение эритроцитов в селезенке, укорочен срок их жизни, повышен гемолиз и часто имеются признаки хронической гипоксии.
Серповидноклеточная анемия наследуется по аутосомно-рецессивному типу. У больных, гетерозиготных по гену серповидно-клеточной анемии (Аа), наряду с серповидными эритроцитами, несущими гемоглобин S, в крови наличествуют и нормальные, несущие гемоглобин А. При этом болезнь менее выражена клинически, протекает легче, а иногда вообще не вызывает симптомов, и серповидные эритроциты выявляются случайно при лабораторном исследовании крови. У гомозигот (аа) по гену серповидноклеточной анемии в крови имеются только серповидные эритроциты, несущие гемоглобин S, болезнь протекает тяжело и заканчивается летальным исходом.
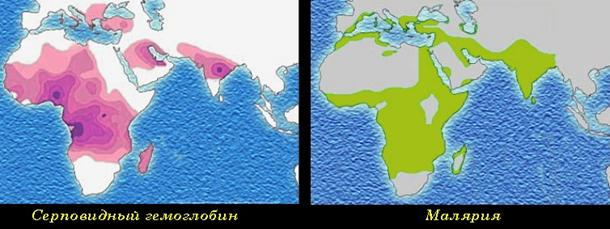
Рис. 34. Строение эритроцитов при серповидноклеточной анемии. Серповидноклеточная анемия весьма распространена в регионах мира, эндемичных по малярии, причем больные серповидноклеточной анемией обладают повышенной (хотя и не абсолютной) врожденной устойчивостью к заражению различными штаммами малярийного плазмодия (рис. 35).
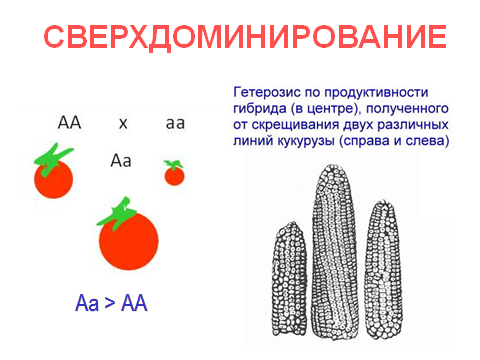
Рис. 35. Распространенность серповидноклеточной анемии и малярии. 3. Сверхдоминирование (рис. 36) – вид взаимодействия аллельных генов, при котором фенотипическое выражение признака у гетерозиготы (Аа) намного ярче, чем у доминантной гомозиготы (АА). Этот вид взаимодействия генов лежит в основе гетерозиса.
Рис. 36. Свехдоминирование. 4. Аллельное исключение (рис. 37) – вид взаимодействия аллельных генов, при котором в части соматических клеток организма, гетерозиготного по данному локусу, активен один аллель, тогда как в других клетках другой. Такой вид взаимодействия аллельных генов наблюдается при наследовании признаков у организмов-мозаиков.
Рис. 37. Аллельное исключение. 5. Кодоминирование – вид взаимодействия аллельных генов, при котором у гетерозиготы отсутствуют доминантно-рецессивные отношения, аллели фенотипически проявляют себя как бы независимо друг от друга. Примером кодоминантного взаимодействия аллелей является наследование IV группы крови у человека.
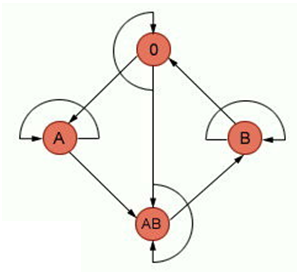
В популяциях людей известны 4 антигенные группы крови человека: А, В, АВ, О и три типа аллелей этого гена: IA, IB, i (табл. 4).
Таблица 4 Особенности наследования групп крови
Генотипы IAIA, IAi имеют эритроциты, содержащие на поверхности только антиген А, а в плазме крови гемагглютинин β (группа крови А или II). Генотипы IBIb, IВi имеют эритроциты, содержащие на поверхности только антиген В, а в плазме крови гемагглютинин α (группа крови В или III). Генотип IAIB имеет эритроциты, несущие оба антигена А и В, а в плазме крови отсутствуют гемагглютинины α и β (группа крови IV или АВ). Это и есть случай кодоминирования (рис. 38). Рис. 38. Наследование IV группы крови. У людей с генотипом ii на поверхности эритроцитов нет антигенов А и В, а в плазме крови присутствуют гемагглютинины α и β. Агглютинин α специфически связывается и осаждает эритроциты с антигеном А, агглютинин β – эритроциты с антигеном В. На этих взаимоотношениях основана система переливания крови. Долгие годы придерживались так называемого закона Отенберга, согласно которому агглютинируют только эритроциты перелитой донорской крови (а не эритроциты реципиента), учитывая, что агглютинины донорской крови разводятся в крови реципиента и не способны агглютинировать его эритроциты. Это обстоятельство разрешало переливать наравне с одногруппной и кровь другой группы, сыворотка которой не агглютинировала эритроциты реципиента. На практике используется следующая схема (рис. 39): реципиенту 0(I) группы допустимо переливать донорскую кровь только 0(I) группы, реципиентам А(II) группы – донорскую кровь А(II) и 0(I) групп, реципиентам В (III) группы – донорскую кровь В (III) и 0(I) групп, реципиентам АВ(IV) группы – донорскую кровь всех четырех групп. Любому реципиенту можно вводить кровь I группы (0), так как ее эритроциты не содержат агглютиногены и не склеиваются, поэтому лиц с I группой крови называют универсальными донорами, но им самим можно вводить кровь только I группы. Кровь от донора IV группы можно переливать только лицам данной группы, но им самим можно переливать кровь всех четырех групп. Людей с IV группой крови называют универсальными реципиентами.
Рис. 39. Схема переливания крови с учетом группы крови. 6. Множественный аллелизм – присутствие в генофонде популяции одновременно 3 и более различных аллелей одного гена. Примером может служить разнообразие окраски глаз человека, разнообразие групп крови. Ген I может быть представлен тремя разными аллелями: IA, IB, i, которые комбинируются в зиготах только попарно. Другим примером является наследование окраски шерсти у кроликов (рис. 40). В популяции кроликов существует четыре аллеля.
Рис. 40. Наследование окраски шерсти в популяции кроликов. Ген А отвечает за наследование темной окраски шерсти и доминируют над всеми другими аллелями. Ген ach обуславливает шиншилловую окраску и по отношению к генам ah и a ведет себя как доминантный. Ген ah отвечает за гималайскую окраску шерсти и доминирует над геном a (белая окраска). Таким образом, А > ach > ah > a (табл. 5).
Таблица 5 Генотипы и фенотипы кроликов
Причиной множественного аллелизма являются случайные изменения структуры гена (мутации), сохраняемые в процессе естественного отбора в генофонде популяции. Многообразие аллелей, рекомбинирующихся при половом размножении, определяет степень генотипического разнообразия среди представителей данного вида, что имеет большое эволюционное значение, повышая жизнеспособность популяций в меняющихся условиях их существования. Кроме эволюционного и экологического значения аллельное состояние генов оказывает большое влияние на функционирование генетического материала. В диплоидных соматических клетках эукариотических организмов большинство генов представлено двумя аллелями, которые совместно влияют на формирование признаков.
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2019-12-15; просмотров: 986; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 52.14.144.75 (0.01 с.) |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||