Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Комплементарность (комбинативное взаимодействие)Содержание книги
Поиск на нашем сайте
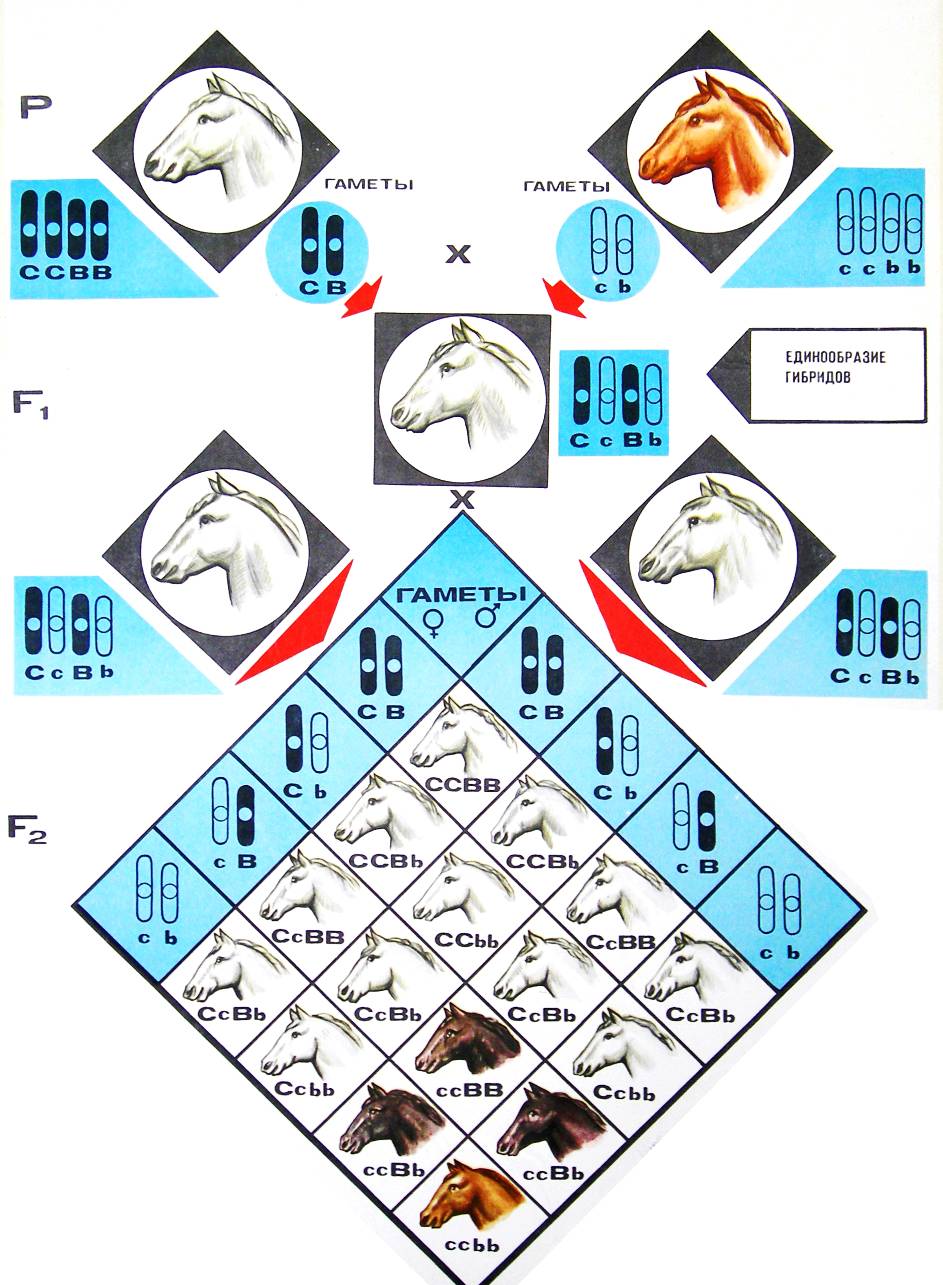
Один из первых примеров взаимодействия неаллельных генов был обнаружен в начале XX в. при анализе наследования формы гребня у кур. Описано четыре разновидности форм гребней, при этом разные породы имеют характерную морфологию гребня (рис. 43):
Рис. 43. Разновидности форм гребней у разных пород кур. · леггорны – листовидный (простой гребень), · виандотты – розовидный (низкий, утолщенный спереди, заостренный сзади и у основания покрытый сосочками), · европейские – гороховидный (невысокий, с тремя продольными пластинками), · малайские – ореховидный, напоминающий поверхность половинки грецкого ореха. В результате скрещиваний кур, имеющих розовидный и гороховидный гребни, в F1 возникает новая форма гребня – ореховидный (из-за взаимодействия генов A и B).
Р: Розовидный Гороховидный ♀ AAbb х ♂ aaBB
G: Ab aB
F1: AaBb – 100% Ореховидный
Скрещивание гибридов F1 дает следующие результаты в F2:
Потомство F2 характеризуется следующими особенностями: 1. Присутствие доминантных аллелей двух генов А и В у 9/16 кур второго поколения ведет к образованию ореховидного гребня. 2. Присутствие гена А в гомо- или гетерозиготном состоянии при рецессивном b дает розовидную форму у 3/16 особей, а гены ааВ – у 3/16 потомства дают гороховидный гребень. 3. Гомозиготы по обоим рецессивным генам ааbb имеют новый фенотип – простой листовидный гребень. Этот признак в последующих скрещиваниях не дает расщепления. Итак, взаимодействие доминантных генов А и В изменяет форму гребня. В этом случае расщепление в дигибридном скрещивании нарушается, однако очевидно, что общее соотношение вариантов 9: 3: 3: 1 сохраняется.
Наследование окраски плодов у перца – 9: 3: 3: 1 (рис. 44).
Рис. 44. Разновидности окраски перца: генотипы R • cc – коричневая окраска, rrC • – желтая; R • C • – красная (комплементарная); rrcc – зеленая. Наследование окраски тела у змей (рис. 45). Во втором гибридном поколении наблюдается расщепление в соотношении 9 : 3: 3: 1. При этом ген А отвечает за синтез красного пигмента, а – отсутствие синтеза красного пигмента; ген В обусловливает полосатый рисунок, b – ромбовидный рисунок.
Рис. 45. Генотипы змей: (а) – ААВВ; (b) – ааВВ; (с) – ААbb; (d) – ааbb. ЭПИСТАЗ (подавление) – ген одной аллельной пары подавляет действие генов другой аллельной пары. Ген, подавляющий действие другого гена, называется эпистатическим геном, ингибитором или супрессором. Подавляемый ген носит название гипостатического.
Различают доминантный и рецессивный эпистаз. При доминантном эпистазе доминантный аллель одного гена (ген-супрессор) подавляет действие доминантной или рецессивной аллели другого гена (гипостатический ген). Ген А (супрессор) подавляет ген В (гипостатический) или ген А (эпистатический) подавляет ген b (гипостатический). При доминантном эпистазе в дигибридном скрещивании происходит расщепление в соотношении 12:3:1 или 13:3. Рис. 46.Расщепление 13:3 в F2 при доминантном эпистазе. Например, при скрещивании 2 белых кур леггорн ААСС и виандотт аасс во втором поколении произойдёт расщепление 13 белых: 3 окрашенных. Ген С подавляет ген А. В отсутствии гена С проявляется ген А, и куры окрашены (рис. 46). У лошадей ген В обусловливает синтез черного пигмента (вороная масть), а ген b – рыжую масть. Ген С – эпистатический ген (серая масть – результат эпистаза), а его рецессивный аллель с не подавляет проявлений гена B и b. Лошади серой масти имеют генотип – 9 •ѕ и 3 bbС•; вороные – 3В•сс, рыжие – 1bbсс. Расщепление 12: 3: 1 (рис. 47).
Рис. 47. Расщепление 12:3:1 в F2 при доминантном эпистазе. В случае рецессивного эпистаза рецессивный эпистатический аллель в гомозиготном состоянии препятствует проявлению доминантной или рецессивной аллели другого гена (аа подавляет ген В или аа подавляет ген b). При рецессивном эпистазе в дигибридном скрещивании происходит расщепление в соотношении 9: 3: 4. Примером рецессивного эпистаза у животных служит окраска шерсти у кроликов (рис. 48). Окраска «агути» у них определяется доминантным геном В, а чёрная окраска его рецессивным геном b. Оба признака проявляются только при наличии доминантного гена А, а его рецессивная аллель (а) подавляет цветность. Рис. 48. Расщепление 9: 3: 4 в F2 при рецессивном эпистазе. Ярким примером рецессивного эпистаза у человека является " Бомбейский феномен ". Так, у человека группы крови АВО контролируются тремя аллелями одного гена IA – вторая группа (синтез антигена А), IB – третья группа крови (синтез антигена В), i – первая группа кровит (синтез антигенов не происходит). В популяции встречается редкий мутантный аллель h независимого гена, который в гомозиготном состоянии подавляет действие аллелей А и В, что приводит к фенотипическому проявлению первой группы крови.
Рис. 49. Генотипы родителей и потомков с фенотипом крови Бомбей. В Индии была описана семья, в которой родители имели вторую и первую группу крови. Оба родителя были гомозиготными по группам крови, и рождение ребенка с четвертой группой крови в их семье вызвало недоумение (рис. 49). Таким образом, "Бомбейский феномен" определяется тем, что в генотипе детей есть ген подавляющий действие генов IА и IВ.
Рис. 50. Часть родословной женщины с фенотипом крови Бомбей. Генетически она имеет группу крови В, но из-за отсутствия антигенов АВ ее кровь отнесена к группе 0. Группа крови АВ0 хорошо исследована биохимически. Известно, что антигены А и В представляют собой карбогидратные группы (углеводные остатки), связанные с жирными кислотами, молекулы которых выступают над поверхностью мембраны эритроцитов. Специфичность этих антигенов обусловлена концевым остатком карбогидратной цепи. Оба антигена происходят из одного и того же предшественника, или Н-вещества, к молекулам которого затем добавляется один или два концевых углеводных остатка сахара. В крайне редких случаях, например, у женщин из Бомбея (Индия) имеется дефектное Н-вещество, которое не связывается с ферментом, добавляющим концевые углеводные остатки. У носителей группы крови 0 это приводит к фенотипу Бомбей. Оказалось, что этот дефект обусловлен рецессивной мутацией h в локусе, не гомологичном локусам А и В. Поэтому при наличии у человека аллелей IA и (или) IB вместе с генотипом hh антигены А и В в крови не обнаруживаются. Вот почему у одной из жительниц Бомбея с генотипом hh определили группу крови 0, хотя ее родители имели группу крови АВ, а сын был носителем аллеля IB (рис. 50). ПОЛИМЕРИЯ – явление, когда несколько неаллельных доминантных генов контролируют развитие одного и того же признака. Такие гены называются полимерными и обозначаются одной и той же буквой, но с другим индексом (например: А1, А2, А3, А4 и т.д.). Различают кумулятивную и некумулятивную полимерию.
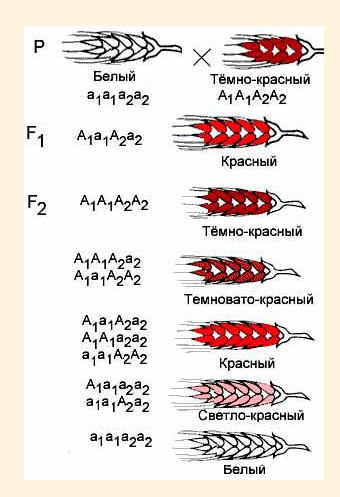
Рис. 51. Некумулятивная полимерия. При некумулятивной полимерии признак проявляется при наличии хотя бы одного из доминантных аллелей полимерных генов. Количество доминантных аллелей не влияет на степень выраженности признака. Впервые некумулятивная полимерия была описана в 1908 г. Нильсоном-Эле при анализе наследования окраски эндосперма зерновок пшеницы (рис. 51). Доминантные неаллельные гены А1 и А2 обуславливают красную окраску зерен пшеницы, их рецессивные аллели а1 и а2 – отсутствие окраски. Действие генов А1 и А2 однозначно. При наличии двух полимерных генов расщепление во втором поколении соответствует 15: 1, при наличии трёх полимерных генов – 63: 1 и т.д.). При кумулятивной полимерии действие полимерных генов суммируется, чем их больше, тем ярче выражен признак. Расщепление во втором поколении будет зависеть от количества полимерных генов (суммарный или аддитивный эффект генов), присутствующих в генотипе. По типу кумулятивной полимерии наследуются многие количественные признаки (у человека: пигментация кожи, рост, вес, уровень интеллекта, скорость протекания биохимических реакций, артериальное давление, содержание сахара в крови; у животных: жирность молока; у растений: яровость, озимость, содержание витаминов, длина колоса у злаков, интенсивность окраски частей растения и др.). Цвет кожи человека определяется взаимодействием нескольких пар генов по типу кумулятивной полимерии, т.е. цвет кожи тем темнее, чем больше доминантных генов в генотипе (рис. 52).

Рис. 52. Кумулятивная полимерия. ЭФФЕКТ ПОЛОЖЕНИЯ ГЕНОВ – влияние расположения генов в хромосоме на проявление их активности. Явление открыто американским генетиком А. Стёртевантом в 1925 году. Наблюдается при структурных перестройках хромосом (транслокациях), в результате которых гены активных зон хромосом (эухроматина) могут переноситься в неактивные зоны (гетерохроматин) и инактивироваться и наоборот. При перестройке, возвращающей эухроматиновый ген из гетерохроматина в любую точку эухроматина, функционирование данного гена восстанавливается. Эффект положения гена можно проследить при наследовании резус фактора (Rh-фактор). Рис. 53. «Эффект положения». В простейшем варианте резус-положительность доминирует над резус-отрицательностью и практически наследование Rh-фактора иммитирует моногогенное наследование. Однако ряд исследований показывает, что система Rh определяется тремя антигенными факторами, которые детермируются тремя тесно сцепленными генами С, Д, Е, локализованными в коротком плече хромосомы 1. Внутри аллельных пар действует полное доминирование, а при взаимодействии между ними «эффект положения». Все это обусловливает разнообразие вариантов резус-антигенов и соответственно резус-несовместимости. Основная роль в Rh-системе принадлежит антигену D. При его наличии на поверхности эритроцитов кровь является резус-положительной. Резус-отрицательный фенотип формируется при отсутствии антигена D. Однако антигены C и E оказывают влияние на резус-несовместимость. Суть «эффекта положения» в следующем: в рассмотренном примере (рис. 53) оба человека имеют одинаковые фенотипы и генотип. Они гетерозиготы по трем парам генов (CcDdEe) и оба резус-положительные, но в антигенном плане их кровь несовместима. У человека, у которого в одной хромосоме порядок расположения генов Cde, а в другой соответственно cDE, будут синтезироваться все три вида антигенов C, D, E и его кровь будет несовместима с кровью второго человека. У второго человека порядок расположения генов внутри хромосомы cDe, а в гомологичной – CdE. Там, где в одной хромосоме расположены доминантные гены С и Е, ген Е выступает в роли супрессора по отношению к гену С. Следовательно, фенотипическое проявление гена С в признак подавлено, антиген С не синтезируется.
|
||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2019-12-15; просмотров: 995; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.216.253.84 (0.009 с.) |
||||||||||||||||||||||||||||||||||||