Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Э.Н. Макаренко, М.В. ПоходенкоСодержание книги
Поиск на нашем сайте
Министерства здравоохранения Российской Федерации
КАФЕДРА БИОЛОГИИ Э.Н. Макаренко, М.В. Походенко ГЕНЕТИКА: ОСНОВЫ НАСЛЕДСТВЕННОСТИ Учебное пособие
Ставрополь 2014
УДК ББК М Макаренко Э.Н. ГЕНЕТИКА: ОСНОВЫ НАСЛЕДСТВЕННОСТИ: учебное пособие для бакалавров / Э.Н. Макаренко, М.В. Походенко – Ставрополь: Изд-во СтГМУ. – 2014 – 78 с. Пособие составлено в соответствии с федеральным государст-венным образовательным стандартом и включает все базовые вопросы общей генетики в разделе «Наследственность». В учебном пособии рассмотрены законы Менделя, хромосомная теория наследственности, классические методы генетики: гибридологический анализ, анализирующее и реципрокное скрещивание. На современном учебно-методическом уровне представлены сведения о видах взаимодействия аллельных и неаллельных генов, специфике проявления гена в признак, генетике пола. Пособие предназначено для бакалавров СтГМУ. Рецензенты: Мануйлов И.М.., профессор кафедры ботаники, зоологии, общей биологии Северо-Кавказского федерального университета, доктор ветеринарных наук. Радцева Г.Л., кандидат медицинских наук, доцент кафедры гистологии СтГМУ. УДК ББК М Рекомендовано к изданию цикловой методической комиссией Ставро-польского государственного медицинского университета. Рекомендовано к печати редакционно-издательским советом СтГМУ. © Ставропольский государственный медицинский университет, 2014 ВВЕДЕНИЕ ГЕНЕТИКА – наука о наследственности и изменчивости. Наследственность – всеобщее свойство живого в виде способности организмов передавать свои признаки и свойства из поколения в поколение. Изменчивость – свойство прямо противоположное наследственности – способность организмов приобретать новые признаки и свойства в процессе индивидуального развития организмов (онтогенеза). 1900 год – год рождения генетики как науки. «Отцом», основоположником генетики считается чешский естествоиспытатель Грегор Мендель (рис.1).
Рис. 1. Грегор Мендель(1822 – 1884). ЗАКОНЫ МЕНДЕЛЯ
В 1865 г. в работе " Опыты над растительными гибридами " Г. Мендель изложил результаты экспериментов по изучению закономерностей наследования отдельных признаков у гороха. Горох обладает следующими характеристиками: 1. Огромное количество сортов с множеством альтернативных признаков (форма, окраска семян и др. (рис. 2)). 2. Самоопыляющееся растение → стойко сохраняет свои сортовые особенности → возможность получения «чистых линий». 3. Легко культивируется в большом количестве. 4. Возможность проведения искусственного перекрестного опыления. 5. Полученные гибриды полностью фертильны.
Рис. 2. Альтернативные признаки садового гороха. Ряд обнаруженных Г. Менделем закономерностей впоследствии стали называться общебиологическими законами наследования или законами Г. Менделя. К сожалению, современники не поняли и не оценили работу Г. Менделя, и она оставалась забытой вплоть до 1900 г., когда подобные результаты получили в разных странах три ученых (рис. 3) – Гуго де Фриз (Голландия), Эрих Чермак (Австрия) и Карл Корренс (Германия). Этот год считается годом переоткрытия законов Г. Менделя и рождения генетики как науки.
Рис. 3. Гуго де Фриз (1848–1935), Эрих Чермак (1871–1962) и Карл Корренс (1864–1933). Для постановки эксперимента при изучении наследования признаков, Г. Менделем был разработан метод гибридологического анализа. Вот основные его свойства: 1) в скрещивании участвуют организмы, принадлежащие к одному виду; 2) изучаемые признаки должны быть взаимоисключающими или контрастными (альтернативными); 3) исходные родительские формы должны быть «чистыми линиями» (гомозиготами) по изучаемым признакам; 4) при изучении закономерностей наследования, необходимо начинать работу с анализа минимума количества признаков, постепенно усложняя эксперимент: родительские особи должны отличаться по одной паре альтернативных признаков → двум парам → небольшому числу пар альтернативных признаков; 5) проводить индивидуальный анализ потомства и при наличии расщепления в поколении необходимо проводить статистический анализ; 6) изучение закономерностей наследования проводится на протяжении нескольких поколений. Таким образом, гибридологический анализ – это система скрещиваний, позволяющая проследить в ряду поколений характер наследования признаков и выявить новообразования. Прежде чем начать основные эксперименты, Г. Мендель провел предварительное исследование экспериментального объекта и тщательно спланировал все эксперименты. Основным принципом исследования была этапность – все внимание сначала концентрировалось на одной переменной, что упрощало анализ, затем Т. Мендель приступал к анализу другой. Все методики строжайше соблюдались, чтобы не вносить искажения в результаты; полученные данные тщательно регистрировались. Г. Мендель провел множество экспериментов и получил достаточное количество данных для обеспечения статистической достоверности результатов. В выборе же экспериментального объекта Г. Менделю, действительно, во многом повезло, поскольку на наследовании отобранных им признаков не сказывались некоторые более сложные закономерности, открытые позднее. Изучая результаты скрещивания растений с альтернативными признаками, Г. Мендель обнаружил, что гибриды первого поколения (F1), полученные с помощью искусственного опыления, не являются промежуточными между двумя родительскими формами, а в большинстве случаев соответствуют одной из них. Тот признак родителя, которым обладали растения первого поколения, Г. Мендель назвал доминантным признаком (от латинского “dominans” – господствующий). От экспериментально полученных гибридов уже путем самоопыления Г. Мендель получил потомство второго поколения (F2) и обнаружил, что эти потомки не являются одинаковыми: часть из них несет признак того родительского растения, который не проявился у гибридов первого поколения. Таким образом, признак, отсутствовавший в поколении F1, вновь проявился в поколении F2. Г. Мендель сделал вывод, что этот признак присутствовал в поколении Fl в скрытом виде. Г. Мендель назвал его рецессивным признаком (от латинского “recessus” – отступление, удаление). Г. Мендель провел целую серию аналогичных опытов с разными парами альтернативных признаков и всякий раз тщательно подсчитывал соотношение растений с доминантными и рецессивными признаками. Во всех случаях анализ показал, что соотношение гибридов с доминантными признаками к потомкам с рецессивными в поколении F2 составляло примерно 3:1. Таблица 1 Условные обозначения Г. Менделя
В третьем поколении (F3), полученном так же путем самоопыления растений из поколения F2, оказалось, что те растения из второго поколения, которые несли рецессивный признак, дали нерасщепляющееся потомство; растения с доминантным признаком частично оказались нерасщепляющимися (константными), а частично дали такое же расщепление, как и гибриды F1 (3 доминантных на 1 рецессивный). Заслуга Г. Менделя состояла в том, что он понял: такие соотношения признаков в потомстве могут быть только следствием существования обособленных и неизменяющихся единиц наследственности – «наследственных факторов», передаваемых с половыми клетками от поколения к поколению. Г. Мендель ввел буквенные обозначения (табл. 1) для доминантного и рецессивного факторов, причем доминантные обозначались большими буквами, а рецессивные – маленькими. Моногибидное скрещивание – родительские особи, взятые для скрещивания, отличаются одной парой альтернативных признаков. Дигибридное скрещивание – организмы, взятые для скрещивания, отличаются двумя парами альтернативных признаков. Закономерности наследования, установленные Г. Менделем при моно- и дигибридном скрещиваниях: § 1 закон – «Закон единообразия гибридов первого поколения»; § 2 закон – «Закон расщепления»; § 3 закон – «Закон независимого наследования (комбинирования) признаков». Закон – «Закон расщепления» В ходе дальнейшего скрещивания гибридов первого поколения, во втором поколении (F2) наблюдается расщепление по генотипу 1:2:1, по фенотипу 3:1.
ЦИТОЛОГИЧЕСКИЕ ОСНОВЫ ЦИТОЛОГИЧЕСКИЕ ОСНОВЫ ЗАКОНАМИ МЕНДЕЛЯ
Как при моно-, так и при дигибридном скрещивании потомство F1 единообразно как по фенотипу, так и по генотипу (проявление первого закона Менделя). В поколении F2 происходит расщепление по каждой паре признаков по фенотипу в соотношении 3:1 (второй закон Менделя). Это свидетельствует об универсальности законов наследования Менделя для признаков, если их определяющие гены расположены в разных парах гомологичных хромосом и наследуются независимо друг от друга. СЦЕПЛЕННОЕ НАСЛЕДОВАНИЕ
Г. Мендель изучил наследование только семи пар признаков у душистого горошка. Его законы подтвердились на самых разных видах организмов, т. е. было признано, что эти законы носят всеобщий характер. Однако позже, в 1906 году У. Бэтсон и Р. Пеннет, проводя скрещивание растений душистого горошка и анализируя наследование формы пыльцы и окраски цветков, обнаружили, что эти признаки не дают независимого распределения в потомстве, гибриды всегда повторяли признаки родительских форм. Постепенно таких исключений из третьего закона Менделя накапливалось все больше. Стало ясно, что принцип независимого распределения в потомстве и свободного комбинирования распространяется не на все гены. Независимое комбинирование признаков (третий закон Менделя) осуществляется при условии, что гены, определяющие эти признаки, находятся в разных парах гомологичных хромосом. Следовательно, у каждого организма число генов, способных независимо комбинироваться в мейозе, ограничено числом хромосом. Однако в организме число генов значительно превышает количество хромосом. Например, у кукурузы изучено более 500 генов, у мухи дрозофилы – более 1 тыс., а у человека – около 2 тыс. генов, тогда как хромосом у них 10, 4 и 23 пары соответственно. Это дало основание предположить, что в каждой хромосоме локализовано множество генов. Экспериментальные исследования явления сцепленного наследования провел выдающийся американский генетик Т. X. Морган со своими сотрудниками. Сцепленным наследованием называется такой вариант наследования признаков, при котором гены, отвечающие за развитие данных признаков, располагаются в одной хромосоме, составляют группу сцепления (рис.7) и наследуются совместно (сцепленно).
Рис. 7. Группы сцепления генов в хромосомах. Если Г. Мендель проводил свои опыты на горохе, то для Т. Моргана основным объектом стала плодовая мушка дрозофила.
Рис. 8. Самка и самец плодовой мушки дрозофилы. Дрозофила каждые две недели при температуре 25° С дает многочисленное потомство. Самец и самка внешне хорошо различимы – у самца брюшко меньше и темнее (рис. 8). Они имеют всего 8 хромосом в диплоидном наборе, достаточно легко размножаются в пробирках на недорогой питательной среде. Томас Морган, как в свое время Грегор Мендель в опытах с горохом, изучал наследование 2 пар признаков: окраска тела и длина крыльев у мушки дрозофилы. Скрещивая мушку дрозофилу с серым телом и нормальными крыльями с мушкой, имеющей темную окраску тела и зачаточные крылья, в первом поколении (F1) Морган получал гибриды, имеющие серое тело и нормальные крылья (ген, определяющий серую окраску брюшка, доминирует над темной окраской, а ген, обусловливающий развитие нормальных крыльев, – над геном недоразвитых). Следовательно, в F1 Томас Морган получил единообразие гибридов первого поколения, как и Грегор Мендель, но в F2 Томас Морган никогда не получал соотношения 9:3:3:1 как в третьем законе Менделя. Тогда для выяснения генотипов особей первого поколения (F1) он провел анализирующее скрещивание. Анализирующее скрещивание проводится с целью установления генотипа исследуемой особи. Для этого исследуемую особь (?) скрещивают с рецессивной гомозиготой (аа). Если в F1 наблюдается расщепление 1:1, то исследуемая особь является по генотипу гетерозиготой – Аа.
Если в F1 наблюдается единообразие, то исследуемая особь является по генотипу гомозиготой – АА или аа.
ОПЫТЫ ТОМАСА МОРГАНА
При проведении анализирующего скрещивания самки (F1) из первого поколения с самцом, имевшим рецессивные признаки, теоретически ожидалось получить потомство с комбинациями этих признаков в соотношении 1:1:1:1. Однако в потомстве явно преобладали особи с признаками родительских форм (41,5% – серые длиннокрылые и 41,5% – черные с зачаточными крыльями), и лишь незначительная часть мушек имела иное, чем у родителей, сочетание признаков (8,5% – черные длиннокрылые и 8,5% – серые с зачаточными крыльями). Такие результаты могли быть получены только в том случае, если гены, отвечающие за окраску тела и форму крыльев, находятся в одной хромосоме. Если гены окраски тела и формы крыльев локализованы в одной хромосоме, то при данном скрещивании должны были получиться две группы особей, повторяющие признаки родительских форм, так как материнский организм должен образовывать гаметы только двух типов – АВ и аb, а отцовский – один тип – аb. Следовательно, в потомстве должны образовываться две группы особей, имеющих генотип АаВb и ааbb. Однако в потомстве появляются особи (пусть и в незначительном количестве) с перекомбинированными признаками, то есть имеющие генотип Ааbb и ааВb. Для того, чтобы объяснить это, необходимо вспомнить механизм образования половых клеток – мейоз. В профазе первого мейотического деления гомологичные хромосомы конъюгируют, и в этот момент между ними может произойти обмен участками. В результате кроссинговера в некоторых клетках происходит обмен участками хромосом между генами А и В, появляются гаметы Аb и аВ, и, как следствие, в потомстве образуются четыре группы фенотипов, как при свободном комбинировании генов. Но, поскольку кроссинговер происходит при образовании небольшой части гамет, числовое соотношение фенотипов не соответствует соотношению 1:1:1:1. АНАЛИЗИРУЮЩЕЕ СКРЕЩИВАНИЕ ОПЫТ №1. ПОЛНОЕ СЦЕПЛЕНИЕ (из F 1 берется самец – Аа Bb)
У дигетерозигот доминантные гены могут располагаться или в одной хромосоме (цис-фаза), или в разных (транс-фаза) (рис. 9).
Рис. 9. 1 – механизм цис-фазы (некроссоверные гаметы); 2 – механизм транс-фазы (некроссоверные гаметы). ОПЫТ №2. НЕПОЛНОЕ СЦЕПЛЕНИЕ. КРОССИНГОВЕР. (из F 1 берется самка – Аа Bb (рис. 10))
Рис. 10. 1 – некроссоверные гаметы; 2 – кроссоверные гаметы. Таким образом, у самки, как и у самца, гены А, B находятся в одной хромосоме. Но у самца они располагаются ближе друг к другу и удерживаются сильнее (ПОЛНОЕ СЦЕПЛЕНИЕ). У самки гены А и В находятся на некотором расстоянии друг от друга, сцеплены более слабо друг с другом, чем у самца, поэтому в 17% наблюдается кроссинговер (НЕПОЛНОЕ СЦЕПЛЕНИЕ). Расстояние между генами измеряется в морганидах – условных единицах, соответствующих проценту кроссоверных гамет или проценту рекомбинантов. Например, расстояние между генами серой окраски тела и длинных крыльев (также черной окраски тела и зачаточных крыльев) у дрозофилы равно 17%, или 17 морганидам. Способ наследования сцепленных генов отличается от наследования генов, локализованных в разных парах гомологичных хромосом. Так, если при независимом комбинировании дигетерозигота АаВb образует четыре типа гамет (АВ, Аb, аВ и аb) в равных количествах, то такая же дигетерозигота в случае сцепления генов А и В образует только два типа гамет – АВ и аb – тоже в равных количествах. Это некроссоверные гаметы – гаметы, в процессе образования которых кроссинговер не произошел. Последние повторяют комбинацию генов в хромосоме родителя.
Нерекомбинанты – гибридные особи, у которых такое же сочетание признаков, как и у родителей. Однако полное сцепление генов наблюдается достаточно редко, и в потомстве обычно бывают представлены все четыре фенотипа. Таким образом, и в этом случае при дигибридном скрещивании образуются новые сочетания признаков – рекомбинантные фенотипы. Было установлено, что кроме обычных гамет в этом случае возникают и другие – Аb и аВ – кроссоверные гаметы, то есть появляются гаметы с новым сочетанием (комбинацией) аллелей, отличающимися от родительской гаметы.
Причиной возникновения новых гамет является обмен участками гомологичных хромосом, или кроссинговер. Если особи с новыми генными комбинациями (рекомбинанты – гибридные особи, имеющие иное сочетание признаков, чем у родителей) встречаются в потомстве реже, чем особи с родительскими фенотипами, то это верный признак сцепленности соответствующих генов. После опытов Томаса Моргана мушка дрозофила стала излюбленным объектом исследования генетиков, потому что: 1) неприхотлива в содержании (легко размножается в лабораторных условиях при температуре 25°Ϲ); 2) выраженный половой диморфизм; 3) очень плодовита (большое число потомков); 4) быстрая смена поколений; 5) только 8 хромосом (небольшое число групп сцепления), но огромное количество альтернативных признаков. Группа сцепления – гены, локализованные в одной хромосоме и наследующиеся совместно. Число групп сцепления соответствует гаплоидному набору хромосом, поскольку группу сцепления составляют две гомологичные хромосомы, в которых локализованы одинаковые гены. У человека количество групп сцепления зависит от пола – женский пол – 23 группы сцепления, у мужчин – 24 группы сцепления, поскольку две половые хромосомы (Х и Y) не являются гомологичными и несут различный набор генов.
ГЕНЕТИКА ПОЛА Пол – совокупность признаков, по которым производится специфическое разделение особей или клеток, основанное на морфологических и физиологических особенностях, позволяющее осуществлять в процессе полового размножения комбинирование в потомках наследственных задатков родителей.
Рис. 11. Первичные и вторичные половые признаки человека. Морфологические и физиологические признаки, по которым производится специфическое разделение особей, называются половыми. Признаки, связанные с формированием и функционированием половых клеток, называются первичными половыми признаками. Это гонады (яичники или семенники), их выводные протоки, добавочные железы полового аппарата, копулятивные органы. Все другие признаки, по которым один пол отличается от другого, получили название вторичных половых признаков (рис.11). К ним относят: характер волосяного покрова, наличие и развитие молочных желез, строение скелета, тип развития подкожной жировой клетчатки, строение трубчатых костей и др. Если такие различия между особями мужского и женского пола существуют, то говорят о половом диморфизме. Половой диморфизм – это анатомические различия между самками и самцами одного и того же биологического вида, помимо половых органов. Обычно он выражается в разном размере (у многих птиц и млекопитающих самцы больше и тяжелее самок), волосяном покрове (яркое проявление – борода у мужчин), окрасе (яркое оперение у самцов птиц, которое крайне важно для привлечения самок) и т.д.
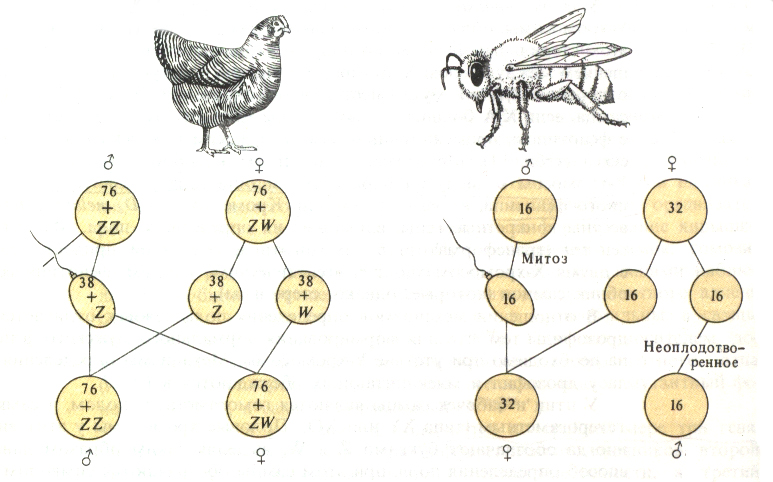
В популяциях животных Начало изучению генотипического определения пола было положено открытием американскими цитологами у насекомых различия в форме, а иногда и в числе хромосом у особей разного пола (Мак-Кланг, 1906, Уилсон, 1906) и классическими опытами немецкого генетика К. Корренса по скрещиванию однодомного и двудомного видов брионии. Уилсон обнаружил, что у клопа Lydaeus turucus самки имеют 14 хромосом (7 пар), как и самцы. Из них 6 пар – одинаковые у обоих полов, а в седьмой паре у самки – две одинаковые хромосомы, а у самца одна хромосома такая же, как соответствующая хромосома самки, а другая маленькая. Пара хромосом, которые у самца и самки разные, получила название гетерохромосомы, или половые хромосомы. У самки две одинаковые половые хромосомы, обозначаемые как Х-хромосомы, у самца одна Х-хромосома, другая – Y-хромосома. Остальные хромосомы, одинаковые у самца и у самки, были названы аутосомами. Таким образом, хромосомная формула у самки названного клопа – 12A + XX, у самца – 12A + XY. В процессе гаметогенеза самка клопа образует один сорт гамет – 7A + X, а самец 2 сорта гамет – 7A + Y или 7A + X. Пол организма с генотипом ХХ называют гомогаметным, так как у него образуются одинаковые гаметы, содержащие только Х-хромосомы, а пол с генотипом XY – гетерогаметным, так как половина гамет содержит Х-, а половина – Y-хромосому. Сочетание XX–XY (рис. 17) определяет пол млекопитающих, человека и некоторых насекомых (клопы, мушка-дрозофила). При гаметогенезе наблюдается типичное менделевское расщепление по половым хромосомам. Каждая яйцеклетка содержит одну Х-хромосому. Сперматозоид содержит одну Y-хромосому или одну Х-хромосому. Пол потомка зависит от того, какой спермий оплодотворит яйцеклетку. При слиянии сперматозоида и яйцеклетки, несущих Х-хромосомы, образуется зигота с двумя Х-хромосомами. В дальнейшем из таких зигот развиваются нормальные самки. Если в зиготу попадают Х- и Y-хромосомы, то в дальнейшем из них развиваются самцы. Сочетание ZZ–ZW (рис. 17) характерно для птиц, бабочек, шелкопряда, рептилий, земноводных. При слиянии сперматозоида и яйцеклетки, несущих Z-хромосомы, образуется зигота с двумя Z-хромосомами. В дальнейшем из таких зигот развиваются нормальные самцы (гомогаметный пол). Если в зиготу попадают Z- и W-хромосомы, то в дальнейшем из них развиваются самки (гетерогаметный пол). В научной литературе вместо символа Z используется Х, имея в виду, что Z-хромосома соответствует Х-хромосоме.
Рис. 17. Гомо- и гетерогаметный пол. Сочетание XX–XО (рис. 18) характерно для некоторых насекомых (кузнечики, моль). Сочетание ХХ характерно для гомогаметного пола, появляется оно при слиянии яйцеклетки и сперматозоида, несущих по Х-хромосоме. Если происходит слияние половой клетки, несущей Х-хромосому, с гаметой, у которой нет ни одной половой хромосомы, образуется сочетание ХО и развиваются организмы гетерогаметного пола.
Рис. 18. Сочетание XX–XО. Совершенно другой механизм определения пола, называемый гаплодиплоидный – сочетание n – 2n широко распространен у пчел и муравьев (рис.19).
Рис. 19. Гаплодиплоидный механизм наследования пола. У этих организмов нет половых хромосом: самки – это диплоидные особи, а самцы (трутни) – гаплоидные. Самки развиваются из оплодотворенных яиц (2n), а из неоплодотворенных путем партеногенеза развиваются трутни (n). Балансовая теория определения пола К. Бриджес в 1919 году обнаружил, что у плодовой мушки дрозофилы на проявление пола оказывает влияние соотношение (баланс) между количеством Х-хромосом и набором аутосом, а Y-хромосома на детерминацию пола влияния не оказывает. У нормальных самок на две Х-хромосомы (2Х) приходится два набора аутосом (2А). Половой индекс в этом случае равен 1 (2Х:2А). У нормальных самцов только одна Х-хромосома. Половой индекс в этом случае равен 0,5 (1Х:2А).
Рис. 20. Самец (а), самка (б) и некоторые ненормальные половые типы дрозофилы: интерсекс (в), сверхсамка (г), сверхсамец (д). В случае неравномерного распределения генетического материала в кариотипе может оказаться на одну Х-хромосому больше (ХХХ или ХХY), а количество аутосом останется в норме. При наличии 3Х хромосом половой индекс будет равен 1,5 (3Х:2А) и разовьются сверхсамки. При наличии 2Х хромосом и 1Y хромосомы половой индекс будет равен 1 (2Х:2А) и появятся нормальные самки, так как Y-хромосома в этом случае на пол влияния не оказывает. При неравномерном распределении генетического материала в кариотипе может оказаться нормальное число половых хромосом, а число наборов аутосом будет кратно увеличено. При кариотипе 2Х+3А половой индекс будет равен 0,67 (2Х:3А) и разовьються интерсексы, а при кариотипе ХY+3А половой индекс будет равен 0,33 (1Х:3А) и разовьются сверхсамцы (бесплодны). Таким образом, следует, что при половом индексе равном 1 развиваются нормальные самки, 0,5 – нормальные самцы, выше 1 – сверхсамки, ниже 0,5 – сверхсамцы, при половом индексе меньше 1, но выше 0,5 – интерсексы (рис. 20). СОДЕРЖАНИЕ
ЛИТЕРАТУРА и ИНТЕРНЕТ-РЕСУРСЫ:
1) Слюсарев А.А. Биология с общей генетикой: учебник. – М.: Альянс, 2011. – 472 с. 2) Никольский В.И. Генетика: учебник, 2010. – 368 с. 3) Курчанов Н.А. Генетика человека. – Санкт-Петербург, 2009. 4) Н.А.Курчанов. Генетика человека с основами общей генетики (уч. пос.). – Санкт-Петербург, СпецЛит, 2009. – 191с. 5) Генетика: учебник для ВУЗов / Под ред. В.И. Иванова. – М.: Академкнига, 2006. 6) Общая генетика (метод.пос.)\ Под ред. С. Г. Инге-Вечтомова. – СПб: Изд-во Н-Л, 2008. – 124 с. 7) Биология (1-3 т.): Д. Тейлор, Н. Грин, У. Стаут. – Москва: Мир, 2008 г. http://900igr.net/kartinki/biologija/Izmenchivost/001-Tema-Nasledstvennaja-izmenchivost.html http://biofile.ru/bio/2604.html http://biofile.ru/bio/6805.html http://www.tepka.ru/biologia10-11/46.html http://900igr.net/kartinki/biologija/Izmenchivost-organizmov/002-Izmenchivost.html
Министерства здравоохранения Российской Федерации
КАФЕДРА БИОЛОГИИ Э.Н. Макаренко, М.В. Походенко ГЕНЕТИКА: ОСНОВЫ НАСЛЕДСТВЕННОСТИ Учебное пособие
Ставрополь 2014
УДК ББК М Макаренко Э.Н. ГЕНЕТИКА: ОСНОВЫ НАСЛЕДСТВЕННОСТИ: учебное пособие для бакалавров / Э.Н. Макаренко, М.В. Походенко – Ставрополь: Изд-во СтГМУ. – 2014 – 78 с. Пособие составлено в соответствии с федеральным государст-венным образовательным стандартом и включает все базовые вопросы общей генетики в разделе «Наследственность». В учебном пособии рассмотрены законы Менделя, хромосомная те
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2019-12-15; просмотров: 548; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.2 (0.01 с.) |










 образуются гаметы:
образуются гаметы: 
 образуются гаметы:
образуются гаметы: