Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Роль антител в элиминации антигеновСодержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
В организме антиген может распространяться с лимфой (лимфогенный путь) и кровью (гематогенный путь) по различным органам и тканям. При этом чаще всего он фильтруется в лимфоузлах, селезенке, а также в лимфоидных скоплениях печени, кишечника и других органов, где вступает в контакт с факторами иммунной защиты. Ответная реакция этих факторов возникает практически немедленно. Первыми вступают в действие факторы врожденного иммунитета, так как эта система не требует длительного времени для активации. Если антиген не был инактивирован или элиминирован в течение 4 ч, включается система приобретенного иммунитета: обеспечивается специфическое распознавание «свой-чужой», вырабатываются факторы регуляции (цитокины) и иммунной защиты (специфические антитела, клоны антигенореактивных лимфоцитов). При этом элиминации подвергаются следующие группы клеток: • терминально дифференцированные лимфоциты, завершившие свою биологическую программу; • активированные лимфоциты, не получившие антигенного стимула; • «изношенные» лимфоциты; • аутореактивные клетки. Естественными факторами, инициирующими апоптоз, яляются глюкокортикоидные гормоны, Fas -лиганд, α-ФНО и другие иммуноцитокины, гранзимы и гранулизин. Апоптотическое уничтожение клеток-мишеней могут активировать Т-киллеры, ЕК с фенотипом CD16-CD56много и Т1-хелперы. 20\ lgM активируют систему комплемента; IgE связывается со специфическими рецепторами на поверхности тучных клеток и базофилов с высвобождением из этих клеток медиаторов аллергии; IgA секретируется в различные жидкости организма, обеспечивая секреторный иммунитет; IgD функционирует в основном в качестве мембранных рецепторов для антигена; в IgG проявляет разнообразные виды активности, в том числе способность проникать через плаценту. Классы иммуноглобулинов. Иммуноглобулины G, IgG Иммуноглобулины G – это мономеры, включающие 4 субкласса (IgGl – 77%; IgG2 – 11%; IgG3 – 9%; IgG4 – 3%), которые отличаются друг от друга по аминокислотному составу и антигенным свойствам. Их содержание в сыворотке крови колеблется от 8 до 16,8 мг/мл. период полураспада составляет 20-28 дней, а синтезируется в течение суток от 13 до 30 мг/кг. На их долю приходится 80% от общего содержания ИГ. Они защищают организм от инфекций. Антитела субклассов IgGl и IgG4 специфически связываются через Fc-фрагменты с возбудителем (иммунное опсонирование), а благодаря Fc- фрагментам взаимодействуют с Fc-рецепторами фагоцитов (макрофагов, полиморфноядерных лейкоцитов), способствуя тем самым фагоцитозу возбудителя. IgG4 участвует в аллергических реакциях и не способен фиксировать комплемент. Антитела класса IgG играют основополагающую роль в гуморальном иммунитете при инфекционных заболеваниях, вызывая гибель возбудителя с участием комплемента и опсонизируя фагоцитарные клетки. Они проникают через плаценту и формируют антиинфекционный иммунитет у новорожденных. Они способны нейтрализовать бактериальные экзотоксины, связывать комплемент, участвовать в реакции преципитации. Иммуноглобулины М, IgM Иммуноглобулины М – это наиболее «ранние» из всех классов ИГ, включающие 2 субкласса: IgMl (65%) и IgM2 (35%). Их концентрация в сыворотке крови колеблется от 0,5 до 1,9 г/л или 6% от общего содержания ИГ. За сутки синтезируется 3-17 мг/кг, а период их полураспада составляет 4-8 суток. Они не проникают через плаценту. IgM появляется у плода и участвует в антиинфекционной защите. Они способны агглютинировать бактерий, нейтрализовать вирусы, активировать комплемент. IgM играют важную роль в элиминации возбудителя из кровеносного русла, в активации фагоцитоза. Значительное повышение концентрации IgM в крови наблюдается при ряде инфекций (малярия, трипаносомозе) как у взрослых, так и у новорожденных. Это показатель внутриутробного заражения возбудителя краснухи, сифилиса, токсоплазмоза, цитомегалии. IgM – это антитела, образующиеся на ранних сроках инфекционного процесса. Они отличаются высокой активностью в реакциях агглютинации, лизиса и связывания эндотоксинов грамотрицательных бактерий.
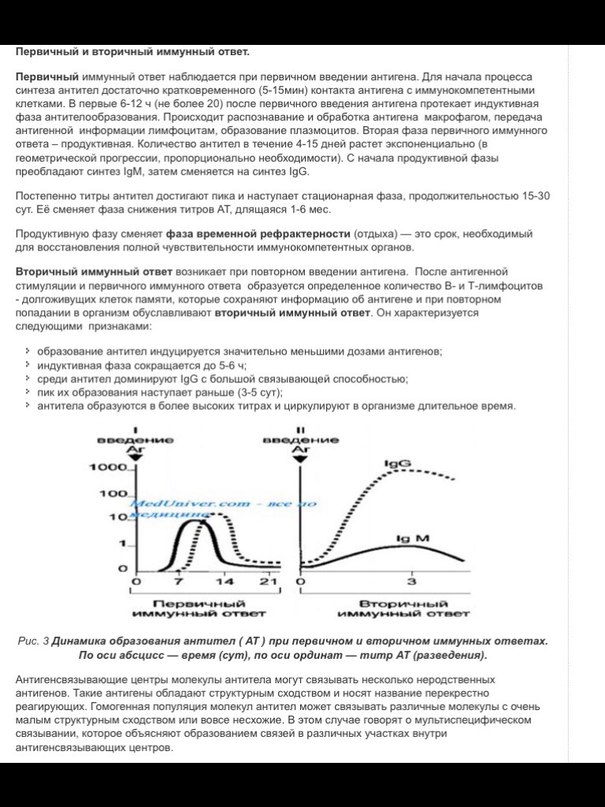
Иммуноглобулины A, IgA Иммуноглобулины А – это секреторные ИГ, включающие 2 субкласса: IgAl (90%) и IgA2 (10%). Содержание IgA в сыворотке крови колеблется от 1.4 до 4.2 г/л или 13% от общего количества ИГ; ежедневно синтезируется от 3 до 50 мкг/кг. Период полураспада антител составляет 4-5 суток. IgA содержится в молоке, молозиве, слюне, в слезном, бронхиальном и желудочно-кишечном секрете, желчи, моче. В состав IgA входит секреторный компонент, состоящий из нескольких полипептидов, который повышает устойчивость IgA к действию ферментов. Это основной вид ИГ, участвующих в местном иммунитете. Они препятствуют прикреплению бактерий к слизистой, нейтрализуют энтеротоксин, активируют фагоцитоз и комплемент. IgA не определяется у новорожденных. В слюне он появляется у детей в возрасте 2 месяца., причем первым обнаруживается секреторный компонент SC. И только позднее полная молекула SigA. Возраст 3 мес. Многими авторами определяется как критический период; этот период особенно важен для диагностики врожденной или транзиторной недостаточности местного иммунитета. Иммуноглобулины Е, IgE Иммуноглобулины Е — это мономеры, содержание которых в сыворотке крови ничтожно мало – 0.00005-0,0003 г/л или 0.002% от общего количества ИГ. За сутки синтезируется 0,02мг/кг, а период их полураспада в сыворотке крови составляет 2-3 дня, а в коже -9-14 дней. К классу IgE относится основная масса аллергических антител – реагинов. Уровень IgE значительно повышается у людей, страдающих аллергией и зараженных гельминтами. IgE связывается с Fc-рецепторами тучных клеток и базофилов. 11ри контакте с аллергеном образуются мостики «IgE-aHTHreH-IgE», что сопровождается поступлением ионов кальция в клетку-мишень, активацией в ней биохимических процессов и выделением БАВ. Вызывающих аллергические реакции немедленного типа. Эозинофильный хемотаксичеекий фактор, выделяемый тучными клетками, способствует аккумуляции эозинофилов и деструкции гельминтов. Предполагается также, что IgE, покрывая паразита, аккумулирует макрофаги благодаря Fc-рецепторам этих клеток. Иммуноглобулины D, IgD Иммуноглобулины D – это мономеры; их содержание в крови составляет 0,03-0.04 г/л или 1% от общего количества ИГ; в сутки их синтезируется от 1 до 5 мг/кг, а период полураспада колеблется в пределах 2-8 дней. IgD участвуют в развитии местного иммунитета, обладают антивирусной активностью, в редких случаях активируют комплемент. Плазматические клетки, секретирующие IgD, локализуются преимущественно в миндалинах и аденоидной ткани. IgD выявляются на B-клетках и отсутствуют на моноцитах, нейтрофилах и Т-лимфоцитах. Полагают, что IgD участвуют в дифференцировке В-клеток, способствуют развитию антиидиотипического ответа, участвуют в аутоиммунных процессах. нная система реагирует на появление во внутренней среде макроорганизма антигена усилением биосинтеза специфических антител. Это достигается размножением клонов антигенспецифических клеток-антителопродуцентов. При этом антиген выступает в роли как пускового, так и селектирующего фактора: преимущественно активируются клоны с наивысшей специфичностью, т.е. наибольшей аффинностью рецепторных молекул Ig. Параллельно с размножением идет процесс дифференцировки В-лимфоцитов. Наблюдаются перестройка в геноме клеток и переключение их биосинтеза с крупной высокоавидной молекулы IgM на более легкие и экономичные высокоаффинные IgG или IgA. Антителопродукция в ответ на антигенный стимул имеет характерную динамику. Ее можно проследить на примере сывороточных Ig (рис. 11.4). Выделяют латентную (индуктивную), логарифмическую, стационарную фазы и фазу снижения. В латентную фазу антителопродукция практически не изменяется и остается на базальном уровне. В этот период происходят переработка и представление антигена иммунокомпетентным клеткам и запуск пролиферации антигенспецифических клонов клетокантителопродуцентов. Ввиду того что клетки делятся дихотомически (т.е. надвое), прирост их количества происходит в логарифмической зависимости и поэтому после первых циклов деления оно изменяется незначительно. Параллельно происходят дифференцировка пре-В-лимфоцитов в зрелые формы и плазматические клетки и переключение синтезируемых изотипов Ig. Во время логарифмической фазы наблюдается интенсивный прирост количества
антигенспецифических В-лимфоцитов, что находит отражение в существенном нарастании титров специфических антител. В стационарной фазе количество специфических антител и синтезирующих их клеток достигает максимума и стабилизируется. Освобождение макроорганизма от антигена устраняет антигенный стимул, поэтому в фазе снижения наблюдается постепенное уменьшение количества клонов специфических антителопродуцентов и титров соответствующих антител. Динамика антителообразования существенно зависит от первичности или вторичности контакта с антигеном. При первичном контакте с антигеном развивается первичный иммунный ответ. Для него характерны длительные латентная и логарифмическая (7-15 сут) фазы. Первые диагностически значимые титры специфических антител регистрируются на 10-14-е сутки от момента иммунизации. Стационарная фаза продолжается 15-30 сут, а фаза снижения - 1-6 мес. В течение первичного иммунного ответа происходят созревание, размножение клонов и дифференцировка антигенспецифических В-лимфоцитов, а также переключение биосинтеза Ig с изотипа M на изотопы G, A или Е. В итоге первичного иммунного реагирования формируются многочисленные клоны антигенспецифических антителопродуцирующих клеток и клеток иммуноло- гической памяти, а во внутренней среде макроорганизма в высоком титре накапливаются специфические IgG и/или IgA. Таким образом обеспечиваются активное противодействие иммунной системы внедрению в макроорганизм антигена и высокая готовность к повторной с ним встрече. Со временем антительный ответ угасает. Элиминация антигена исключает новое стимулирование к клонообразованию, а появившиеся ранее плазматические клетки имеют короткую продолжительность жизни. Вместе с тем В-лимфоциты иммунологической памяти надолго остаются циркулировать в организме. Повторный контакт иммунной системы с тем же антигеном ведет к формированию вторичного иммунного ответа (см. рис. 11.4). Его латентная фаза значительно укорочена, а логарифмическая фаза отличается более интенсивной динамикой прироста и более высокими титрами специфических антител. Для стационарной фазы и фазы снижения свойственна затяжная динамика (несколько месяцев или даже лет). При вторичном иммунном ответе организм сразу же в подавляющем большинстве синтезирует IgG. Это обусловлено подготовленностью иммунной системы к повторной встрече с антигеном за счет формирования иммунологической памяти (см. раздел 11.5): многочисленные клоны антигенспецифических В-лимфоцитов, оставшиеся после первичного иммунного реагирования, быстро размножаются и интенсивно включаются в процесс антителогенеза. Для развития гуморального иммунитета слизистых оболочек характерны те же процессы и динамика антителообразования. Однако в данном случае в слизистых оболочках в подавляющем большинстве созревают и размножаются В-лимфоциты, продуцирующие полимерные молекулы IgA. Явление интенсивного антителообразования при повторном контакте с антигеном широко используется в практических целях, например вакцинопрофилактике. Для создания и поддержания иммунитета на высоком защитном уровне схемы вакцинации предусматривают многократное введение антигена для формирования и поддержания иммунологической памяти (см. главу 14). Этот же феномен используют при получении высокоактивных лечебных и диагностических иммунных сывороток (гипериммунных). Для этого животным или донорам производят многократные введения препаратов антигена по специальной схеме. Динамика и интенсивность антителообразования в значительной степени зависят от иммуногенности антигена: дозы, способа и кратности его введения, а также от состояния макроорганизма. Попытка повторного введения антигена в латентной фазе может привести к иммунологическому параличу - иммунологической неотвечаемости на антиген в течение определенного периода времени. 22. Метод парных сывороток -это такая методика в лабораторных исследованиях, когда кровь берут у одного исследуемого или пациента дважды через определенный промежуток времени и о заболевании судят не по абсолютным показателям, а по динамике, нарастанию этих показателей, что говорит о том, что в организме пациента идет активный процесс или же, в случае отсутствия такой динамики, процесс уже закончен (т. е, возможно уже переболел, иммунитет уже сработал, поэтому показатели номинально большие, но человек уже поправился 23. B-лимфоци́ты (B-клетки, от bursa fabricii птиц, где впервые были обнаружены) — функциональный тип лимфоцитов, играющих важную роль в обеспечении гуморального иммунитета. У эмбрионов человека и других млекопитающих B-лимфоциты образуются в печени и костном мозге из стволовых клеток, а у взрослыхмлекопитающих — в красном костном мозге. При контакте с антигеном или стимуляции со стороны T-клеток некоторые B-лимфоциты трансформируются в плазматические клетки, способные к продукции антител. Другие активированные B-лимфоциты превращаются в B-клетки памяти. Дифференцировка В-лимфоцитов Лимфоциты происходят от плюрипотентных стволовых клеток, дающих также начало всем клеткам крови. Дифференцировка стволовых клеток крови по эритроидному, миелоидному либо лимфоидному пути зависит от микроокружения (в случае птиц дифференцировка стволовых клеток в В-лимфоциты происходит в фабрициевой сумке, у млекопитающих в костном мозге, где также происходит дифференцировка по миелоидному и эритроидному пути). Дифференцировка В-лимфоцитов условно делится на две стадии — антигеннезависимую (в которую происходит перестройка генов иммуноглобулинов и их экспрессия) и антигензависимую (при которой происходит активация, пролиферация идифференцировка в плазматические клетки). · Пре-В-Клетки-предшественники не синтезируют тяжёлых и лёгких цепей, содержат зародышевые H и L гены, но содержат антигенный маркер, общий с зрелыми пре-В-клетками. · Ранние пре-В-клетки — D-J перестройки в Н генах. · Поздние пре-В-клетки — V-DJ перестройки в Н генах. · Большие пре-В-клетки Н-гены VDJ-перестроены; в цитоплазме имеются тяжёлые цепи класса μ. · Малые пре-В-клетки — V-J перестройки в L генах; в цитоплазме имеются тяжёлые цепи класса μ. · Малые незрелые В-клетки — L гены VJ-перестроены; синтезируют Н и L- цепи; на мембране расположены иммуноглобулины. · Зрелые В-клетки- начало синтеза IgD. В-клетки поступают из костного мозга во вторичные лимфоидные органы (селезёнку и лимфатические узлы), где происходит их дальнейшее созревание, антиген-презентирование, пролиферация и дифференцировка в плазматические клетки и В-клетки памяти. В-клетки B-лимфоцит окарашенный гемотоксилином и эозином под оптическим микроскопом. Экспрессия всеми В-клетками мембранных иммуноглобулинов позволяет осуществляться клональному отбору под действием антигена. При созревании, антиген-стимулировании и пролиферации существенно меняется набор маркеров В-клеток. По мере созревания В-клетки переключаются от синтеза IgM и IgD на синтез IgG, IgA, IgE (при этом у клеток сохраняется способность синтезировать также IgM и IgD- вплоть до трёх классов одновременно). При переключении синтеза изотипов антигенная специфичность антител сохраняется. Различают: · Собственно В-клетки (ещё называемые «наивными» В-лимфоцитами) - неактивированные В-лимфоциты, не контактировавшие с антигеном. Не содержат тельца Голла, в цитоплазме рассеяны монорибосомы. Полиспецифичны и имеют слабое сродство к многим антигенам. · В-клетки памяти - активированые В-лимфоциты, посредством кооперации с Т-клетками вновь перешедшие в стадию малых лимфоцитов. Являются долгоживущим клоном В-клеток, обеспечивая быстрый иммунный ответ и выработку большого количества иммуноглобулинов при повторном введении того же антигена. Названы клетками памяти, так как позволяют иммунной системе «помнить» антиген на протяжении многих лет после прекращения действия антигена. В-клетки памяти обеспечивают долговременный иммунитет. · Плазматические клетки - являются последним этапом дифференцировки активированных В-клеток, провзаимодействовавшими с антигеном. В отличие от остальных В-клеток несут мало мембранных антител и способны секретировать растворимые антитела. Являются большими клетками с эксцентрично расположенным ядром и развитым синтетическим аппаратом - шероховатыйэндоплазматический ретикулум занимает почти всю цитоплазму, также развит и аппарат Гольджи. Являются короткоживущими клетками (2-3 дня) и быстро элиминируются при отсутствии антигена, вызвавшего иммунный ответ. Маркеры В-клеток Характерной особенностью В-клеток является наличие поверхностных мембрано-связанных антител, относящихся к классам IgM и IgD. В комплексе с с другими поверхностными молекулами иммуноглобулины формируют антиген-распознающий рецептивный комплекс, ответственный за узнавание антигена. Также на поверхности В-лимфоцитов расположены антигены МНС класса II, важные в кооперации с Т-клетками, также на некоторых клонах В- лимфоцитов присутствует CD5 маркер, общий с Т-клетками. Рецепторы C3b компонента комплемента(Cr1, CD35) и C3d (Cr2,CD21) имеют определённую роль в активации В-клеток. Следует отметить, что маркеры CD19, CD20 и CD22 используются для идентификации В-лимфоцитов. Также на поверхности В-лимфоцитов обнаружены Fc рецепторы. Активация В-клеток Антиген-презентирующая клетка (макрофаги, клетки Купфера, фолликулярные дендритные клетки, интердигитальные дендритные клетки и т. д.) вскоре после процессирования патогена выносит эпитопы на поверхность клетки при помощи МНС II, делая их доступными для Т-клеток. Т-хелпер при помощи Т-клеточного рецептора распознаёт комплекс эпитоп-МНС. Активированный Т-хелпер выделяет цитокины, усиливающие антиген-презентирующую функцию, а также цитокины, активирующих В-лимфоцит - индукторы активации и пролиферации. В-лимфоциты присоединяются при помощи мембрано-связанных антител, выступающих в роли рецепторов, к "своему" антигену и в зависимости от получаемых от Т-хелпера сигналов пролиферируют и дифференцируются в плазматическую клетку, синтезирующую антитела, либо перерождается в В-клетку памяти. При этом от качества и количества антигена будет зависеть исход данной трёхклеточной системы взаимодействия. Данный механизм справедлив для полипептидных антигенов, относительно неустойчивых к фагоцитарному процессингу - т. н. тимус-зависимых антигенов. Для тимус-независимых антигенов (обладающих высокой полимерностью с часто повторяющимися эпитопами, относительно устойчивых к фагоцитарному перевариванию и обладающих свойствами митогена) участия Т-хелпера не требуется - активация В-лимфоцитов происходит по тимус-независимому пути, В-лимфоциты связываются с данными антигенами, а за счёт их собственной митогенной активности будет происходить пролиферация В-лимфоцитов и активация. Роль В-лимфоцитов в презентации антигена В-клетки способны поглощать свои мембранные иммуноглобулины вместе со связанным ими антигеном, а затем презентировать фрагменты антигена в комплексе с молекулами МНС класса II. При низкой концентрации антигена и при вторичном иммунном ответе В-клетки могут выполнять роль основных антигенпрезентирующих клеток. В1- и В2-клетки К популяции В-клеток можно отнести две субпопуляции: B-2 и В-1 клетки. В-1— это большая группа В-клеток у человека и мышей. Они могут составлять около 5% от общей популяции B-клеток. Такие клетки появляются в течение эмбрионального периода. На своей поверхности они экспрессируют IgM и небольшое количество (или вовсе не экспрессируют) IgD. Маркером этих клеток является CD5. Однако он не является обязательным компонентом клеточной поверхности. У животных В-1 клетки находятся в лимфоузлах и селезёнке. В эмбриональном периоде В-1 клетки появляются из стволовых клеток костного мозга. B1–лимфоциты поддерживают свою физиологическую регенерацию в течение всей жизни из отдельной клетки–предшественницы, пул которой у взрослых не пополняется за счёт общей стволовой кроветворной клетки костного мозга. Эта отдельная клетка–предшественница отселяется из кроветворной ткани на свою анатомическую территорию — в брюшную и плевральную полости — ещё в эмбриональном периоде. Итак, место обитания B1–лимфоцитов — прибарьерные полости. B1–лимфоциты значительно отличаются от B2–лимфоцитов по антигенраспознавательным способностям продуцируемых антител. Антитела, синтезированные B1–лимфоцитами, не имеют значительного разнообразия вариабельных участков молекул иммуноглобулинов, но, напротив, ограничены в репертуаре распознаваемых антигенов, и эти антигены — наиболее распространённые соединения клеточных стенок бактерий. Все B1–лимфоциты — как бы один не слишком специализированный, но определённо ориентированный (антибактериальный) клон. Антитела, продуцируемые B1–лимфоцитами, почти исключительно IgM, переключение классов иммуноглобулинов в B1–лимфоцитах не «предусмотрено». Таким образом, B1–лимфоциты — «отряд» противобактериальных «пограничников» в прибарьерных полостях, предназначенных для быстрой реакции на «просачивающиеся» через барьеры инфекционные микроорганизмы из числа широко распространённых. В сыворотке крови здорового человека преобладающая часть иммуноглобулинов — продукт синтеза как раз B1–лимфоцитами, т.е. это относительно полиспецифичные иммуноглобулины антибактериального назначения.
|
||||||
|
|
Последнее изменение этой страницы: 2017-02-10; просмотров: 540; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.217.161.27 (0.011 с.) |

 Рис. 11.4. Динамика антителообразования при первичном (I) и вторичном (II) иммунном ответах. Фазы антителообразования: а - латентная; б - логарифмического роста; в - стационарная; г - снижения
Рис. 11.4. Динамика антителообразования при первичном (I) и вторичном (II) иммунном ответах. Фазы антителообразования: а - латентная; б - логарифмического роста; в - стационарная; г - снижения