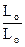Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Механіка м'язового скороченняСодержание книги
Поиск на нашем сайте
За сучасними уявленнями усі м'язи розділяються на три групи: 1. кістякові, 2. гладкі, 3. м'язи серця (міокард). Міокард являє собою основну масу серця. При його скороченні зменшується обсяг порожнин серця (передсердь, шлуночків), підвищується тиск, який забезпечує безперервний плин крові. Гладкі м'язи входять до складу стінок внутрішніх органів (шлунку, кишечнику, судин, сечового міхура й т.п.). При напрузі цих м'язів відбувається зміна форми органів, необхідна для здійснення біологічних процесів (руху крові, лімфи, сили, їжі й ін.). Кістякові м'язи складаються з великої кількості паралельно розташованих волокон, що представляють собою високоспеціалізовані клітини. Як і будь-які інші клітини, вони відмежовані мембраною й містять органоїди: ядра, саркоплазматичний ретикулум, мітохондрії й ін. Особливе значення для виконання скорочувальної функції належить тонким (діаметром 1-2 мкм) ниткам - міофібрилам, які тягнуться від одного кінця волокна до іншого. У першому наближенні можна вважати, що довжина міофібрил дорівнює довжині м'яза. Кількість цих органоїдів у клітині досить велика й може досягати 1000-2000.
Рис. 10 За допомогою мікроскопічних спостережень у міофібрилах виявляються поперечно розташовані зони А, I, Н, Z (див. рис. 10). Наявність поперечної зчерченості дала підставу називати кістякові м'язи поперечно-смугастими. Відзначимо, що в м'язі серця також виявляється поперечна зчерченість.
Рис. 11 У міофібрилах можна виділити послідовно повторюваний елемент - саркомер, обмежений Z - дисками, вихідна довжина якого зменшується в процесі скорочення. За сучасними уявленнями саме саркомер є елементарною моторною одиницею, а механізм його скорочення лежить в основі діяльності всіх кістякових м'язів. При зменшенні довжини саркомеру (відстані між Z-дисками) змінюються розміри й взаєморозташування зон А, I, H. Як показують результати електронно-мікроскопічного спостереження, ці зсуви пояснюються взаємодією більш тонких ниток - протофібрил (рис.11). Виявлені протофібрили двох типів - тонкі, що складаються із білка актину, і товсті, що містять білок - міозин. Зіставлення результатів електронної й оптичної мікроскопії дозволяє пояснити поперечну зчерченість: Н - відстань між кінцями актинових ниток у центрі саркомера, I - проміжок між кінцями міозинових ниток поруч розташованих саркомерів. В основі теоретичного пояснення скорочення саркомера лежить модель ковзних ниток.
Рис. 12 При подразненні м'язів нервовими імпульсами або електричним струмом із саркоплазматичного ретикулума вивільнюються іони кальцію, які сприяють формуванню зв'язків (містків) між міозиновими й актиновими нитками (рис. 12). Актиноміозиновий комплекс, що утворюється, має ферментативну активність і розщеплює універсальне джерело енергії - молекули АТФ. Звільнена при цьому енергія передається на кальцієві містки, які працюють у циклічному режимі. Містки забезпечують рух актинових ниток до центру смуги в Н-зону, в результаті чого Z-перегородки наближаються до кінців міозинових ниток. Цей молекулярний механізм створює зусилля, що сприяє переміщенню протофібрил м'язового волокна один щодо одного доти, поки кінці міозинових ниток не досягнуть Z-дисків (див. рис. 12). За існуючими оцінками при розщепленні молекули АТФ один місток за повний цикл роботи протягом 1 мс розвиває зусилля 1012 H. Оскільки саркомери розташовуються в міофібрилі послідовно, вихідну довжину м'яза, можна виразити Lo = nlo, де lo - довжина саркомера до скорочення, n - кількість саркомерів. У процесі зменшення відстані між Z - дисками на = lo - l довжина всієї міофібрили змінюється на L = Lo - L = nlo - nl = n(lo - l) = n . Відношення величини вкорочення м'яза до її вихідної довжини Lo називається відносним укороченням: = L/Lo = n /nlo = /lo. Як видно з наведеного співвідношення, відносне вкорочення м'яза можна розрахувати у такий спосіб. Скориставшись геометричними розмірами, визначеними з мікроскопічних спостережень (довжина lo = 2.00 - 2.25 мкм, розмір зони А a = 1.6 мкм), можна визначити абсолютне = lo - a = 0.6 - 0.65 мкм і відносне вкорочення саркомера = /lo =0.27 - 0.29 = 27 - 29%. Аналогічні розрахунки показують, що максимальне відносне вкорочення для більшості м'язів становить величину порядку 30%. Отже, для одержання значних переміщень L = необхідно, щоб рух забезпечувався м'язами з великою вихідною довжиною. Визначимо залежність зміни довжини м'яза від часу. Очевидно, швидкість скорочення в початковий момент часу найбільша, потім зменшується й, коли вкорочення досягає максимуму L, стає рівною нулю. Така залежність може бути описана наступним виразом: V = k(L - X), де Х - укорочення в момент часу, L - найбільше вкорочення при t і k - коефіцієнт пропорційності, що залежить від прикладеного до м'яза навантаження. Швидкість скорочення є першою похідною від укорочення за часом: V = dx/dt = k(L - X). Вирішуючи це диференціальне рівняння, знайдемо вираз, що описує вкорочення за часом: X = L (1 - e- kt) або X = (1 - e- k t), де - відносне вкорочення, Lo - вихідна довжина м'яза. Швидкість скорочення визначиться як перша похідна від укорочення: V = dx/dt = e-k t = Vo e-k t На рисунку 13 наведені графіки залежності вкорочення Х і швидкості V від часу t.
Рис. 13 Якщо міофібрили розташовуються уздовж осі м'яза, кількість містків між протофібрилами залежить від площі поперечного переріза м'яза S, і тому силу скорочення можна виразити як: F = = f S, де S - площа розтину, f - питома сила скорочення, яку розвивають містки на одиницю розтину. Остання величина для кістякових м'язів людини становить 30 - 40 Н/м2.
Рис. 14 Залежно від умов стимуляції спостерігається різний характер діяльності м'язів: одиночне скорочення, зубчастий і гладкий тетанус. Коли на м'яз впливає одиночний електричний імпульс, сила, яку він розвиває, змінюється з часом за законом, наведеним на рисунку 14а. Таке скорочення називається одиночним і закінчується в момент t1. Якщо впливати серією електричних подразників із частотою f = 1/t2 причому t2 < t1, то новий електричний імпульс буде дратувати м'яз у момент, коли він ще повністю не розслабився й не відновив вихідну довжину. У цьому випадку сила, що розвивається, не досягає нуля, а змінюється за законом, наведеним на рисунку 14b. Такий режим скорочення називається зубчастим тетанусом.
Якщо дратувати м'яз із ще більшою частотою f = 1/t3, він взагалі не встигає розслаблюватися й створювана сила описується гладкою кривою. Подібне скорочення називається гладким тетанусом (рис. 15). При гладкому тетанусі напруга м'яза перевищує напругу одиночного скорочення у 1,5 - 2 рази. Це пов'язане з тим, що в м'язі є пружні елементи (позафібрилярні компоненти сполучної тканини), які при активному укороченні м'яза скручуються на зразок пружини й перешкоджають розвитку напруги в скорочувальних елементах за час t1. При переході одиночних скорочень у тетанус послідовно розташовані пружні елементи деформуються й не виявляють такої протидії, як при одиночному скороченні. Необхідно відзначити, що цей механізм відіграє вирішальну роль у розслабленні м'язів. Після скорочення іони кальцію видаляються із цитоплазми в саркоплазматичний ретикулум, актиноміозиновый комплекс розпадається й за рахунок внутрішнього напруження деформованих пружних елементів протофібрили повертаються у вихідний стан. Величину механічної роботи в процесі м'язового скорочення можна виразити: А = F L = fSL, де F - м'язова сила (за умови, що вона постійна), L - абсолютне вкорочення, дорівнює L = Lo - L. Уважаючи в першому наближенні, що м'яз може бути представлений циліндром, помножимо й поділимо величину роботи на вихідну довжину Lo: А = f S L Добуток SLo виражає вихідний об'єм Vo, а = L/Lo - відносне вкорочення м'яза. Тоді робота м'яза А = fVo. Якщо представити об'єм через щільність і масу м'яза Vo = M/, то: А = (f/) Відношення питомої сили до щільності для даної м'язової тканини є постійною величиною k = f/, і тоді остаточно робота при м'язовому скороченні: А = k .
Рис. 16
Із цієї рівності випливає, що механічна робота залежитьвід маси й відносного вкорочення м'яза. Величина сили, яку розвивають протофібрили, визначається довжиною саркомера. Це пояснюється тим, що в процесі ковзання ниток, по-перше, змінюється кількість містків, що утворюються між протофібрилами, а по-друге, - при мінімальній довжині саркомера міозинові нитки впираються в Z - перегородки (рис. 16). На рисунку 17 наведена експериментальна залежність між відносною силою й довжиною саркомера. F (%) = (F/Fm)100 (де F - сила, що розвивається, а Fm - максимальна сила скорочення). При довжині саркомера, l > 3.65 мкм між протофібрилами не можуть утворюватися містки, і тому сила дорівнює нулю. У міру зменшення відстані між активновими й міозиновими нитками утворюється все більша кількість містків і тому сила зростає. Найбільшу силу саркомер розвиває, коли l лежить у діапазоні 2 - 2,25 мкм. Якщо відстань l стає менше 2,05 мкм, актинові нитки перекриваються й зусилля F падає.
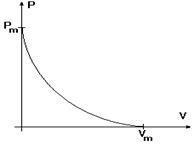
Рис. 17 При відстані між Z-дисками менше 1,65 мкм кінці міозинових ниток упираються в перегородки, вони деформуються, порушується їхня просторова структура, й тому сила, що розвивається, стає рівною 0 (при l = 1,05 мкм). РІВНЯННЯ ХІЛА Рівняння Хіла є основним рівнянням динаміки м'язового скорочення й описує взаємозв'язок між силою й швидкістю вкорочення: (Р + а)(V + b) = b (Pm + a) = const
Рис. 18 Тут позначено: Р - сила, V - швидкість, яку розвиває м'яз, a, b, Pm - постійні величини. Із цієї рівності випливає висновок, що між силою й швидкістю існує обернено пропорційна залежність. Якщо швидкість V = 0, то підстановка цього значення приводить до співвідношення P = Pm, тобто в цьому випадку м'яз розвиває максимальну силу, рівну Pm. Якщо сила скорочення дорівнює нулю, швидкість досягає максимальної величини Vm = bPm/a. При інших значеннях сил і швидкостей взаємозв'язок між ними описується гіперболічною залежністю, що наведена на рисунку18. Детальне дослідження рівняння Хіла дозволяє виявити три режими діяльності м'язів: ізометричний, ізотонічний і ауксотонічний. При ізометричній напрузі довжина м'яза не змінюється (Lo = const), укорочення L= 0, і швидкість, як перша похідна від укорочення з часом, дорівнює нулю (V = dx/dt = 0). З рівняння Хіла випливає, що в цьому режимі сила, яку розвиває м'яз, досягає максимального значення Pm. На практиці ізометричний режим зустрічається в тому випадку, коли м'яз працює проти навантаження, що перевищує за величиною значення Pm. Іншим важливим прикладом ізометричного режиму є скорочення міокарда, коли закриті клапани серця й відбувається об'ємний стиск крові, що перебуває в замкненому просторі шлуночків. В ізотонічному режимі м'яз розвиває постійну силу, за величиною меншу Pm. Граничним випадком є таке скорочення, коли Р=0, а швидкість досягає максимального значення Vm. В організмі ізотонічне скорочення практично не зустрічається, тому що м'язи працюють проти змінних навантажень. У реальних умовах найпоширеніший ауксотонічний режим, при якому як сила, так і швидкість змінюються з часом.
|
||||
|
|
Последнее изменение этой страницы: 2017-02-17; просмотров: 189; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.217.98.175 (0.007 с.) |






 Рис. 15
Рис. 15