Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Досягнення сучасної біофізикиСтр 1 из 15Следующая ⇒
Лекція 1 Тема: „Біофізика як наука”
План 1. Визначення та основні завдання біофізики. 2. Розділи біофізичної науки. 3. Історичний розвиток біофізики. 4. Сучасні досягнення та наукові напрямки.
Біофізика – це наука, яка вивчає фізичні й фізико-хімічні процеси, що протікають у біосистемах на різних рівнях організації і є основою фізіологічних актів. Виникнення біофізики відбулося на межі фізики, математики, хімії й біології. Основними об'єктами вивчення є молекули, клітини та живі організми. Живі організми – відкрита, саморегулююча, що самовідтворюється й розвивається гетерогенна система, найважливішими функціональними речовинами в якій є біополімери: білки й нуклеїнові кислоти складної атомно-молекулярної будови. Завдання біофізики: 1. Розкриття загальних закономірностей поведінки відкритих нерівновагих систем. Теоретичне обґрунтування термодинамічних (т/д) основ життя. 2. Наукове тлумачення явищ індивідуального й еволюційного розвитку, саморегуляції й самовідтворення. 3. З'ясування зв'язків між будовою й функціональними властивостями біополімерів й інших біологічно активних речовин. 4. Створення й теоретичне обґрунтування фізико-хімічних методів дослідження біооб'єктів. 5. Фізичне тлумачення великого комплексу функціональних явищ (генерація й розповсюдження нервового імпульсу, м'язове скорочення, рецепція, фотосинтез й ін.) Розділи біофізики: 1. Молекулярна - вивчає будову і фізико-хімічні властивості молекул. 2. Біофізика клітини - вивчає особливості будови й функціонування клітинних і тканинних систем. 3. Біофізика складних систем - вивчає кінетику біопроцесів, поведінку з часом різноманітних процесів, що властиві живий матерії й термодинаміку біологічних систем. Однак, історично сформоване коло проблем, який займається Б., ширше. До Б. відносяться: вивчення впливу фізичних факторів на організм (вібрація, прискорення, невагомість); дослідження біологічної дії іонізуючого випромінювання, яке у зв'язку з важливістю й актуальністю цього питання стало предметом радіобіології -спеціальної науки, що виділилася з Б. Фізичний аналіз діяльності органів почуттів, у першу чергу, оптики ока, аналіз роботи органів руху, подиху, кровообігу як фізичних систем, питання міцності й еластичності тканин (біомеханіка) — істотні, історично сформовані розділи Б. Важливе значення має й розробка фізичних методів дослідження біологічних систем - від макромолекул до цілого організму, без яких неможливо сучасне біологічне дослідження.
Окремі дослідження біофізичного характеру можна простежити з 17 в. У цей період були зроблені спроби застосувати поняття, сформульовані для фізики й хімії, для аналізу біологічних явищ. Французький учений Р. Декарт розглядав людське тіло як складну машину. Він опублікував ряд робіт з дослідження органів почуттів — біоакустиці і оптиці. Послідовник Декарта - італійський учений Дж. А. Бореллі намагався пояснювати рух живих істот чисто фізичними закономірностями. Л. Ейлер, професор Петербурзького університету, уперше математично описав рух крові по судинах. М. В. Ломоносов висунув в 1756 одну з перших гіпотез кольорового зору. Могутнім поштовхом до фізико-хімічних досліджень явищ життя послужили досвіди італійського вченого Л. Гальвані, що довів наявність «тваринної електрики». В 2-й половині 19 в. німецькі вчені Г. Гельмгольц і В. Вундт сформулювали основні закономірності фізіологічної акустики й фізіологічної оптики. Німецький лікар Ю. Р. Майер, спостерігаючи насичення киснем гемоглобіну в крові людини в тропічному й помірному кліматі, сформулював закон збереження енергії. М. Гельмгольц і М. Рубнер продовжили дослідження цього закону на живих організмах. Роботами німецьких учених Г. Гельмгольца, Э. Дюбуа-Реймона, Д. Бернштейна й ряду ін. були закладені основи положень про механізм виникнення електричних потенціалів у тканинах і поширення збудження по нерву. Значення іонного складу й реакції середовища в житті клітин і тканин було з'ясовано в роботах американського дослідника Ж. Леба, німецьких учених В. Нернста й Р. Гебера. У Росії И. М. Сєченов наприкінці 19 в. досліджував фізичні закономірності розчинення газів у крові й біомеханіку рухів. К. А. Тімірязєв вивчав фотосинтетичну активність окремих ділянок сонячного спектра у зв'язку з розподілом енергії в ньому й особливостями спектра поглинання хлорофілу (1903). А. Ф. Самойлов описав акустичні властивості середнього вуха. П. П. Лазарєву належить заслуга в розвитку іонної теорії збудження (1916). М. Н. Шатерніков використав термодинамічні положення в дослідженнях енергетичного балансу організмів (1910-1920). В 1905-1915 роках виконані класичні дослідження Н. К. Кольцова щодо ролі фізико-хімічних факторів (поверхневого натягу, концентрації водневих іонів, катіонів) у житті клітини. Цей етап передісторії Б., що охоплює період до 20 р. 20 в., характерний появою окремих робіт з використанням ідей і методів фізики й фізичної хімії при дослідженні руху, слухового й зорового апаратів, фотосинтезу, механізму генерації електрорушійної сили в нерві й м'язі, значення іонного середовища для життєдіяльності клітин і тканин.
Після Жовтневої революції склалися сприятливі умови для розвитку Б. у СРСР. В 1919 П. П. Лазарєв створив у Москві інститут біологічної фізики, де вели роботи з іонної теорії збудження, кінетики реакцій, що йдуть під дією світла, досліджували спектри поглинання й флуоресценції біологічних об'єктів, а також процеси первинної дії на організм різних чинників зовнішнього середовища. Пізніше такі інститути були створені й в інших країнах. В 20-і рр. Кольцов сформулював концепцію про молекулярну структуру гена й матричний механізм передачі спадкоємної інформації й синтезу макромолекул. В 20-30-і рр. вийшов ряд книг, які зробили глибокий вплив на наступний розвиток Б. у СРСР: «Біосфера» В. И. Вернадського (1926), „Теоретична біологія” Э. С. Бауера (1935), «Фізико-хімічні основи біології» Д. Л. Рубінштейна (1932), «Організація клітини» Н. К. Кольцова (1936), «Реакція живої речовини на зовнішні впливи» Д. Н. Насонова й В. Я. Александрова (1940). У ці роки йшло поступове формування бази для біофізичних досліджень, розроблялися нові методи, росло технічне оснащення лабораторій. Після 2-й світової війни в СРСР і провідних капіталістичних країнах у результаті величезного розмаху досліджень з фізики та хімії, виникнення потужної приладобудівної промисловості й різкого збільшення фінансування біологічних досліджень починається бурхливий розвиток Б. Формування окремих областей Б. Молекулярна Б. досліджує механізм біологічних явищ із погляду взаємодії атомів і молекул, іонів і радикалів. У завдання цього розділу входить вивчення просторової структури, будови, фізико-хімічних властивостей біологічних систем на молекулярному рівні. Ця проблематика тісно пов'язана з біохімією, що особливо яскраво видно на прикладі вивчення будови біологічно важливих макромолекул, з'ясування просторової структури яких вимагає біофізичного підходу й вирішується методом рентгеноструктурного аналізу. Останній був успішно використаний щодо розшифровки простих біологічних молекул (в 20-х рр. в Англії В. Астбери вдалося частково розшифрувати структуру молекули целюлози). Роботи зі структури білка були початі в 30-х рр. англійським ученим Дж. Берналом. У 1954 англійські дослідники Дж. Кендрю й М. Перуц знайшли метод розрахунку просторового розташування атомів у молекулі білка. Це призвело до розрахунку структури міоглобіну й гемоглобіну, що дозволило розкрити механізм виникнення серповидної клітинної анемії й глибше зрозуміти природу активного центру білкової молекули. Роботи з вивчення просторової структури білків велися в СРСР на фізичному факультеті МГУ, в інституті біофизики АН СРСР й інших установах. Дослідження структури фібрилярних білків (колагену, фиброїну, шовку) показали наявність регулярної структури з груп амінокислот, що періодично чергуються. Побудовано статистичну теорію редуплікації (подвоєння) дезоксирібонуклеїнової кислоти (ДНК). До 1968 року визначена структура близько 200 білків. Поряд з вивченням будови окремих молекул більші успіхи досягнуті в дослідженні молекулярних комплексів - ультраструктур, що створюють функціональні одиниці клітини.
Дослідження з молекулярної Б. тісно пов'язані з біохімією, генетикою й цитологією, молекулярною біологією. Значне місце в молекулярній Б. займає проблема збуджених станів молекул у біологічних системах; такі молекули здобувають високу хімічну активність. Найбільш вивчені збуджені стани, що виникають на первинній стадії фотобіологічних процесів — фотосинтезу, зорута біолюмінесценції. Оригінальним напрямком у вітчизняній Б. можна вважати вивчення надслабкого ультрафіолетового світіння біологічних систем (мітогенетичного випромінювання, А. Г. Гурвич, 1923—1948). В 30-і рр. Г. М. Франк і С. Ф. Родіонов розробили фізичний метод виявлення надслабких світінь біологічних об'єктів. Успіхи в розробці методів реєстрації надслабких світлових потоків за допомогою фотоелектронних помножувачів привели в 50-х р. 20 ст. до відкриття надслабких світіння ряду тваринних і рослинних об'єктів у видимій області спектра. Був показаний зв'язок цього світіння з рекомбінацією вільних радикалів. А. Н. Тереніним зі співробітниками були досліджені механізми елементарних фотофізичних процесів за участю пігментів, зазначена роль станів молекул, відкритий механізм міграції енергії в них при фотохімічних реакціях, вивчений механізм люмінесценції білків (1950—1965). А. А. Красновський відкрив і досліджував реакцію оборотного фотохімічного відновлення хлорофілу і його аналогів (1949-1960). Ці роботи сприяли розвитку біологічної фотохімії. В одному з важливих розділів Б. розглядається перетворення енергії в живих організмах, починаючи з перетворення й міграції енергії на молекулярному рівні й кінчаючи енергетичним балансом цілого організму (біоенергетика). Дослідження взаємної трансформації хімічної й механічної енергії при скороченні м'язового волокна, молекулярні механізми руху війок і джгутиків у найпростіших, руху протоплазми й клітинних органел стали предметом вивчення механохімії, що перебуває на стику біохімії й молекулярної Б. В 1938 у роботі радянських учених В. А. Енгельгардта й М. Н. Любимової, що вивчали механізм м'язового скорочення, була вперше продемонстрована наявність прямого зв'язку між механічними й хімічними процесами. Надалі ці роботи були розвинені американським ученим А. Сент-Дьєрд'ї.
Традиційний розділ Б. — вивчення фізико-хімічних властивостей клітини й проникності біологічних мембран для різних речовин. Все більшого значення набувають проблеми моделювання штучних мембран й активного транспорту іонів. Одним із прикладів практичного застосування знань, отриманих у цій області Б., біохімією й фізіологією, є створення штучної нирки. Важливою проблемою Б. є вивчення біоелектричних явищ. У цій області Б. тісно пов'язана з фізіологією (біоелектричні потенціали). Дослідження показали, що між зовнішнім і внутрішнім середовищем кожної живої клітини підтримується різниця потенціалів близько 0,1 в. Її джерело — створюваний клітиною іонний градієнт між зовнішнім і внутрішньоклітинним середовищем. Ці дані послужили основою для створення мембранної теорії генерації потенціалів у клітині, висунутої на початку століття німецьким ученим Д. Бернштейном й експериментально обґрунтованої в 50—60-і рр. роботами англійських учених А. Ходжкіна, А. Хакслі й Б. Каца, що вивчали зміну проникності мембрани нервового волокна й іонні потоки в нерві при збудженні (мембранна теорія збудження). Значне місце займають також дослідження інших фізико-хімічних властивостей клітин - в'язкості, оптичних властивостей, їхніх змін при різних фізіологічних станах та інших впливах. Біофізичні закономірності, властиві організму в цілому, розглядаються у відповідних розділах біоенергетики (вивчення механізму тепловіддачі, теплопродукції, швидкості охолодження при різних умовах і т.п.). Б. процесів керування нерозривно пов'язана з кібернетикою та біомеханікою. Створенню систем керування, з'ясуванню принципів керування рухами тварин і людини поклали початок дослідження радянського вченого Н. А. Бернштейна. Він першим приступився до вивчення зворотних зв'язків у біологічних системах (1934). Вивчення біомеханіки рухів (ходьба, біг, трудові рухи й ін.), подиху й кровообігу має виняткову важливість у зв'язку з питаннями фізіології праці й спорту, космічними польотами, а також для вивчення причин серцевих і судинних захворювань і створення апаратів штучного дихання й кровообігу.
Питання до самоконтролю 1. Біофізика як наука: предмет та завдання, розділи. 2. Формування різних напрямків дослідження в біофізиці. 3. Розвиток Біофізики в Україні та світі.
Лекція 2 Вступ Важко повірити, що 50-60 років тому існування мембран на поверхні живої клітини ставилося під сумнів, а структури, спостережувані під електронним мікроскопом, ніяк не зв'язували із численними функціями живої клітини. Особливо гарячі суперечки стосувалися мембранного механізму виникнення й поширення біопотенціалів. Очевидно, із цієї причини розділ клітинної біофізики, присвячений мембранному електрогенезу, виявився найбільше фундаментально дослідженим і досяг високого ступеня наукової глибини і ясності.
Слід зазначити, що для дослідження мембран потрібна була інтеграція знань багатьох областей природних наук. Щоб зрозуміти, як функціонують мембрани, треба знати такі теми як "Електрика" (фізика), "Ліпіди й білки" (органічна хімія), уміти вирішувати хоча б найпростіші диференціальні рівняння (математика). Структура мембран Перша модель будови біологічних мембран була запропонована в 1902 році. Овертон помітив, що через мембрани найкраще проникають речовини, добре розчинні в ліпідах, і на підставі цього припустив, що біологічні мембрани складаються з тонкого шару фосфоліпідів. Насправді, на поверхні роздягнула полярного й неполярного середовищ (наприклад, води й повітря) молекули фосфоліпідів утворять мономолекулярний (одномолекулярний) шар. Їх полярні "голови" занурені в полярне середовище, а неполярні "хвости" орієнтовані убік неполярного середовища. Тому можна було припустити, що біологічні мембрани побудовані з моношару ліпідів. В 1925 році Гортер і Грендел показали, що площа моношару ліпідів, екстрагованих з мембран еритроцитів, у два рази більше сумарної площі еритроцитів. Гортер і Грендел екстрагували ліпіди з гемолізованих еритроцитів ацетоном, потім випарювали розчин на поверхні води й вимірювали площу мономолекулярної плівки ліпідів, що утворилася. На основі результатів цих досліджень було зроблене припущення, що ліпіди в мембрані розташовуються у вигляді бімолекулярного шару. Це припущення підтвердили дослідження електричних параметрів біологічних мембран (Коул і Кертис, 1935 рік): високий електричний опір ~107 Ом · м2 і більша електроємність ~ 0,5 ·10-2 Ф/м2. Біологічну мембрану можна розглядати як електричний конденсатор. Провідникові пластини конденсатора утворюють електроліти зовнішнього й внутрішнього розчинів (позаклітинного й цитоплазми). Провідники розділені ліпідними бішаром. Ліпіди - діелектрики з діелектричною проникністю Ємність плаского конденсатору становить Питома ємність (на одиницю площини)
Звідси можна знайти відстань між пластинами конденсатору, відповідну у нашому випадку товщині мембрани,
Це якраз відповідає за порядком величини товщині неполярної частини бімолекулярного шару ліпідів, складених визначеним чином. Разом з тим були експериментальні дані, які свідчили про те, що біологічна мембрана містить у своєму складі й білкові молекули. Наприклад, при вимірі поверхневого натягу клітинних мембран було виявлено, що вимірювані значення коефіцієнта поверхневого натягу істотно ближче до коефіцієнта поверхневого натягу на границі розділу білок-вода (близько 0,1 Н/см), ніж на границі розділу ліпід-вода (близько 10 Н/см). Ці протиріччя експериментальних результатів були усунуті Даниеллі й Девсоном, що запропонували в 1935 році так називану "бутербродну" модель будови біологічних мембран, що з деякими несуттєвими змінами протрималася в мембранології майже 40 років. Відповідно до цієї моделі, мембрана - тришарова: вона утворена двома розташованими по краях шарами білкових молекул, з ліпідним бішаром посередині; утвориться щось подібне до бутерброда - ліпіди, на зразок масла, між двома "скибами" білка. Однак у міру накопичення експериментальних даних довелося зрештою відмовитися й від "бутербродної" моделі будови біологічних мембран. Величезну роль у розвитку уявлень про будову біологічних мембран зіграло все більше проникнення в біологію фізичних методів дослідження. Більшу інформацію про структуру мембран, про взаємне розташування атомів мембранних молекул дає рентгеноструктурний аналіз, заснований на дифракції короткохвильових рентгенівських променів на атомах. Рентгеноструктурний аналіз дозволяє виявляти впорядкованість у розташуванні атомів і визначати параметри впорядкованих структур (наприклад, відстані між кристалографічними площинами). Дослідження дифракції рентгенівських променів підтвердили відносно впорядковане розташування ліпідних молекул у мембрані (було показане існування подвійного молекулярного шару з більш-менш паралельно розташованими жирнокислотними хвостами), дали можливість точно визначити відстань між полярною головою ліпідної молекули й метильною групою наприкінці вуглеводневого ланцюга. Найбільш вражаючі результати були отримані в електронно-мікроскопічних дослідженнях. Як відомо, світловий мікроскоп не дозволяє розглянути деталі об'єкта менше приблизно половини довжини хвилі світла (близько 200 нм). У світловому мікроскопі можна розглянути окремі клітини, однак він зовсім не придатний для вивчення біологічних мембран, товщина яких в 20 разів менше межі дозволу світлового мікроскопа. Роздільна здатність мікроскопа обмежена явищем дифракції. Тому, чим менше довжина хвилі в порівнянні з деталями досліджуваного об'єкта, тим менше викривлення. Межа дозволу Z пропорційний довжині хвилі l. Електронам, розігнаним до більших швидкостей, теж властиві хвильові властивості, у тому числі явище дифракції, однак при досить більших швидкостях довжина хвилі де Бройля досить мала й відповідно мала межа дозволу електронного мікроскопа. Так, якщо електрони прискорюються в електричному полі з напругою U = 105 B, швидкість їх досягає 106 м/с (e - заряд електрона, m - маса електрона), а довжина хвилі зменшується та межа дозволу Z становить не тисячні частки нанометрів, а біля одного десятої нанометра (Z ї 0,1 нм), що, однак, уже дозволяє розглянути окремі деталі будови біологічних мембран. До методів вивчення динаміки мембран, що дають можливість досліджувати їх, не руйнуючи, ставляться флюоресцентный метод і методи радіоспектроскопії - електронний парамагнітний резонанс (ЭПР) і ядерний магнітний резонанс (ЯМР). Ці методи дають відомості про рух і взаємодію мембранних молекул й окремих частин молекули. Було з'ясовано, що при фізіологічних умовах ліпідні молекули перебувають у рідкому агрегатному стані. Метод ЭПР показав, що не вся поверхня біологічної мембрани покрита білками. Так, наприклад, більше половини поверхні мембрани кишкової палички утворена полярними головами ліпідів. Сукупність результатів, отриманих фізичними й хімічними методами дослідження, дала можливість запропонувати нову модель будови біологічних мембран - рідинно-мозаїчну (Сингер і Никольсон, 1972 рік) [1 - 3]. Згідно Сингеру й Никольсону, структурну основу біологічної мембрани становить подвійний шар фосфоліпідів, інкрустований білками, подібно тому, як інкрустація кольоровими камінчиками й скельцями створює мозаїчну картину. При цьому розрізняють поверхневі (або периферичні) і інтегральні білки (рис. 2.1).
Рис. 2.1. Будова мембрани за Сингером та Нікольсоном. Мембранні білки можуть пронизувати бислой наскрізь (інтегральний білок - 1), примикати до бішару (периферичний білок - 2) або поринати в нього. Багато білків мембрани є глікопротеїнами (3), а мембраноутворюючі ліпіди - гліколіпідами (4). На схемі також показаний: холестерол (5); вуглевод (6); елементи цитоскелету (7).
Ліпіди перебувають при фізіологічних умовах у рідкому агрегатному стані, це дозволяє зрівняти мембрану з фосфоліпідним морем, по якому плавають білкові "айсберги". Одним з підтверджень рідинно-мозаїчної моделі є й той факт, що, як установлено хімічним аналізом, у різних мембранах співвідношення між змістом білків і фосфоліпідів сильно коливається: кількість білків у миєліновій мембрані в 2,5 рази менше, ніж ліпідів, а в мітохондріях, навпроти, білків в 2,5 рази більше, ніж ліпідів, у той час як, згідно "бутербродної" моделі, співвідношення кількості білків і ліпідів у всіх мембранах повинне бути однаковим. Крім фосфоліпідів і білків у біологічних мембранах містяться й інші хімічні сполуки. У мембранах тваринних клітин багато холестерину (у порівнянній кількості з фосфоліпідами й білками). Є в мембранах й інші речовини, наприклад, гликоліпіди, глікопротеїди. Рідинно-мозаїчна модель будови мембрани в цей час загальноприйнята. Однак, як усяка модель, вона дає досить спрощену й схематичну картину будови мембрани. Зокрема, виявлено, що білкові "айсберги" не завжди вільно плавають у ліпідному морі, а можуть бути "заякорені" на внутрішні (цитоплазматичні) структури клітини. До таких структур відносяться мікрофіламенти й мікротрубочки. Мікротрубочки - порожні циліндри діаметром близько 300 нм із особливого білка тубуліну відіграють, очевидно, важливу роль у функціонуванні клітини. Фізико-хімічні властивості мембрани (рідинність, флексибільність, здатність до агрегації тощо) визначаються, зокрема, її фосфоліпідним складом. Фосфоліпіди є структурними компонентами біологічних мембран (рис. 2.2). Один кінець їх молекули містить два ліпофільні жирно-кислотні ланцюги, другий — гідрофільну іонізовану групу (залишок фосфорної кислоти та основи). Ці молекули впорядковані у фосфоліпідний бішар, який має вигляд сендвіча. На рисунку круглі «головки» представляють іонізовану частину молекули, а «хвости» — ацильні залишки жирних кислот. Центральна частина мембрани містить ліпофільні молекули холестерину, який ущільнює її. Завдяки цьому феномену взаємодія між ліпофільними компонентами мембрани посилюється, мембрана ефективно утримує внутрішній вміст клітини, не даючи йому «вилитися».
Рис. 2.2. Хімічна структура фосфоліпіду: а – полярна (гідрофільна) «головка» (залишок фосфорної кислоти та основи); б, в – неполярні (ліпофільні) ланцюги жирних кислот; г – схематичне зображення молекули фосфоліпіду. (Додатково. Одним із найбільш метаболічно активних фосфоліпідів є фосфатидилінозитол. Під впливом фермента фосфоліпази С від нього відщеплюється інозитолтрифосфат, який стимулює вихід кальцію в клітину із внутрішньоклітинних депо, що може спричинити скорочення гладких м’язів судин. Тому в лікуванні серцево-судинних захворювань особливого значення надають пригніченню цього шляху обміну фосфатидилінозитолу, що ефективно здійснюється антагоністами кальцію. Крім того, посилений метаболізм фосфатидилінозитолу відіграє важливу роль в патогенезі маніакальних станів. Солі літію, ефективні при біполярних розладах, пригнічують цей біохімічний процес. Оскільки полярні (іонізовані) головки фосфоліпідів знаходяться на зовнішній та внутрішній поверхні мембранного «сендвіча», то з ними активно взаємодіють позитивно та негативно заряджені іони. Товща ліпідного матриксу мембрани пронизана білковими молекулами, які виконують у клітині різноманітні функції (див. рис. 2), зокрема рецепторні, ферментативні, канальні тощо. Деякі білки, змінюючи свою динамічну структуру під дією відповідних фармакологічно активних сполук, виконують канальну функцію, формуючи мембранні пори для іонів та малих молекул. Зовнішня та внутрішня гідрофільна поверхня мембрани вкрита водною кристалічною решіткою. Перш ніж молекула лікарського засобу зв’яжеться зі специфічним мембранним рецептором або пройде через мембрану, водна кристалічна решітка має бути частково зруйнована. Речовини перетинають мембранний бар’єр за допомогою активного їх транспорту всередину клітини. Цей процес здійснюється за допомогою спеціальних білків-насосів — натрієвого, кальцієвого, хлорного, протонного та ін. У складі зовнішнього листка плазматичної мембрани знаходяться також вуглеводи, які ковалентно приєднані до білків (глікопротеїди) та ліпідів (гліколіпіди). Вважають, що основна їх функція — участь у процесах міжклітинного розпізнавання, рецепції біологічно активних молекул та забезпеченні імунологічних властивостей клітин).
Модельні ліпідні мембрани Ліпосоми (штучні мембранні частки) використаються в медицині з метою спрямованої доставки ліків. Вікова мрія фармакологів про інструмент, за допомогою якого можна впливати на окрему клітину, близька до здійснення. Плоскі бішарові фосфоліпідні мембрани (БЛМ) - тип модельних мембран. Такі мембрани одержують на маленьких отворах діаметром близько 1 мм, у пластинці із пластику (наприклад, фторопласта), зануреної у водне середовище. На отвір наносять краплю розчину ліпіду (у спирті, хлороформі, гептані або інших розчинниках). Розчинник дифундує з розчину у воду й на отворі залишається плівка ліпіду. Ця плівка спонтанно тоншає доти, поки не утвориться бімолекулярний шар товщиною близько 6 нм. Зайвий ліпід збирається у вигляді ободка - торуса в країв отвору. Плоскі ліпідні мембрани, поряд з ліпосомами, широко використаються як моделі для вивчення електричних властивостей мембрани, їхньої проникності для різних речовин і для інших наукових досліджень. Модельні мембрани моделюють ряд функцій біологічних мембран, у тому числі бар'єрну: наприклад, селективність проникності - гарну проникність для води, погану для іонів. Можна моделювати регульований транспорт, впливаючи на модельну мембрану білками - іонофорами [3, 4], зміною температури, хімічного складу навколишнього середовища, електричним полем [5]. Можна при цьому спостерігати зміну іонної проникності модельних мембран. Як показано фізичними методами дослідження - дилатометрією (виміром коефіцієнта об'ємного розширення) і калориметрією (виміром теплоємності), рентгеноструктурного аналізу й ін. - ліпідна частина біологічних мембран при певних температурах зазнає фазовий перехід першого роду. Згідно даним рентгеноструктурного аналізу, радіоспектроскопії, флюоресцентного аналізу, інфрачервоної спектроскопії й інших фізичних досліджень у фосфоліпідній мембрані при зниженні температури відбувається перехід з рідкокристалічного до гель-стану, що умовно іноді називають твердокристалічним. У гель-стані молекули розташовані ще більш упорядковано, чим у рідко кристалічному. Всі гідрофобні вуглеводневі хвости фосфоліпідних молекул у гель-фазі повністю витягнуті строго паралельно один одному (мають повністю трансконформацию). У рідкому кристалі за рахунок теплового руху можливі структурні переходи: хвости молекул вигинаються, їхня паралельність один одному в окремих місцях порушується, особливо сильно в середині мембрани. Товщина мембрани тому в гель-фазі більше, ніж у рідкому кристалі. Однак при переході із твердого в рідкокристалічний стан, об'єм трохи збільшується, тому що значно збільшується площа мембрани, що доводиться на одну молекулу (від 0,48 нм2 до 0,58 нм2). Для нормального функціонування мембрана повинна бути в рідкокристалічному стані. Тому в живих системах при тривалому зниженні температури навколишнього середовища спостерігається адаптаційна зміна хімічного складу мембран, що забезпечує зниження температури фазового переходу. Температура фазового переходу знижується при збільшенні числа ненасичених зв'язків у жирно-кислотних хвостах. У хвості молекули може бути до чотирьох ненасичених зв'язків. Залежно від хімічного складу ліпідних мембран температура фазового переходу гель-рідкий кристал може мінятися від - 20?З (для мембран ненасичених ліпідів) до + 60?З (для насичених ліпідів). Збільшення числа ненасичених ліпідів у мембрані при зниженні температури перебування спостерігається в мікроорганізмів, рослинних і тварин клітин. Цікавий приклад пристосування клітинних мембран до температурних умов - зміна температури фазового переходу (за рахунок зміни хімічного складу мембранних ліпідів) ноги полярного оленя. Температура уздовж ноги полярного оленя від копита до тулуба взимку може мінятися від - 20°С до + 30°С. Клітинні мембрани в дистальної частини ноги оленя містять більше ненасичених фосфоліпідів. Очевидно, первинний механізм кріопошкоджень (ушкоджень при охолодженні) біологічних мембран пов'язаний з фазовим переходом у гель-стан. Тому біологічні мембрани містять велика кількість холестерину, що зменшує зміни в мембрані, що супроводжують фазовий перехід. Молекули холестерину, розташовуючись між фосфоліпідними молекулами, упорядковують бішар у рідкому й розупорядковують у твердому стані й у такий спосіб зменшують розходження рідко кристалічної і гель-структур. У деяких мікроорганізмів біологічні мембрани перебувають при температурах, що лише небагато перевищують температуру фазових переходів ліпідів. Мембрана містить десятки різних ліпідів, яким відповідають різні температури фазового переходу, у тому числі близькі до фізіологічного. При зниженнях температури в мембрані відбуваються фазові перетворення в ліпідному бішарі. У роботах В.Ф. Антонова й співробітників [5] доведено, що при фазових переходах з гель- у рідкокристалічний стан і назад у ліпідному бішарі утворяться наскрізні канали радіусом 1 - 3 нм, по яких через мембрану можуть переноситися іони й низькомолекулярні речовини. Внаслідок цього при температурі фазового переходу різко збільшується іонна провідність мембрани. Збільшення іонної провідності мембран може врятувати клітину від кріопошкоджень за рахунок збільшення виходу із клітини води й солей - привести до порушення її бар'єрної функції, що перешкоджає кристалізації води усередині клітини. Підвищення іонної провідності мембран при фазовому переході, можливо, дозволяє підтримати на високому рівні метаболічний обмін деяких організмів. Великий інтерес представляє цей ефект для пояснення термо- і хеморецепції. Відомо, що перенос іонів через мембрану лежить в основі формування біопотенциалів, зміна іонної провідності обумовлює нервовий імпульс. Не виключено, що нервовий імпульс, що свідчить про зниження або підвищення температури, утворюється за рахунок зміни іонної проникності ліпідного бішару при фазовому переході мембранних ліпідів нерва. Очевидно, і деякі види хеморецепції можуть бути пов'язані з фазовим переходом мембранних ліпідів, оскільки фазовий перехід може бути викликаний не тільки зміною температури, але й зміною хімічного складу навколишнього середовища. Наприклад, доведено, що при даній температурі фазовий перехід з рідкокристалічного стану в гель-стан може бути викликаний збільшенням концентрації Ca2+ у фізіологічному діапазоні від 1 до 10 мм у водяному розчині, що оточує мембрану. Питання до самоконтролю 1. Основні функції біологічних мембран. 2. Різні моделі структури мембран. 3. Модельні ліпідні мембрани та фазові переходи у мембранах. 4. Модельні ліпідні мембрани.
Лекція 3 План Ендо- та екзоцитоз. Ендоцитоз і екзоцитоз
Ендоцитоз і екзоцитоз - це два активні процеси, за допомогою яких різні матеріали транспортуються через мембрану або в клітини (ендоцитоз), або із клітин (екзоцитоз). 1. Фагоцитоз - поглинання твердих часток. Спеціалізовані клітини, що здійснюють фагоцитоз, називаються фагоцитами (рис. 4).
Рис. 4. Макрофаг, що фагоцитує дві червоні кров'яні клітини. 2. Піноцитоз - поглинання рідкого матеріалу (розчин, колоїдний розчин, суспензія). Часто при цьому утворюються дуже дрібні пухирці (мікропіноцитоз).
Типи проникнення речовин у клітину через мембрани. Молекули проходять через мембрани завдяки трьом різним процесам: простій дифузії, полегшеній дифузії, активному транспорту (рис. 5).
Рис. 5. Типи транспорту крізь мембрану.
Проста дифузія - приклад пасивного транспорту. ЇЇ напрямок визначається тільки різницею концентрацій речовини по обидва боки мембрани (градієнтом концентрації). Шляхом простої дифузії в клітину проникають неполярні (гідрофобні) речовини, розчинні в ліпідах і дрібні незаряджені молекули (наприклад, вода). Більшість речовин, необхідних клітинам, переноситься через мембрану за допомогою занурених у неї транспортних білків (білків-переносників). Усі транспортні білки, очевидно, утворюють безперервний білковий прохід через мембрану. Розрізняють дві основні форми транспорту за допомогою переносників: полегшена дифузія й активний транспорт. Полегшена дифузія обумовлена градієнтом концентрації, і молекули рухаються відповідно до цього градієнту. Однак якщо молекула заряджена, то на її транспорт впливає як градієнт концентрації, так і загальний електричний градієнт поперёк мембрани (мембранний потенціал). Активний транспорт - це перенос розчинених речовин проти градієнта концентрації або електрохімічного градієнта з використанням енергії АТФ. Енергія потрібна тому, що речовина повинна рухатися всупереч своєму природньому прагненню дифундувати в протилежному напрямку. Деякі транспортні білки переносять одну розчинена речовина через мембрану (уніпорт). Інші функціонують як котранспортні системи (рис. 6), у яких перенос однієї розчиненої речовини залежить від одночасного або послідовного переносу другої речовини. Друга речовина може транспортуватися в тому ж напрямку (симпорт) або в протилежному (антипорт).
Рис. 6. Котранспортні системи. Na-K насос
|
|||||||||
|
|
Последнее изменение этой страницы: 2017-02-17; просмотров: 282; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.222.240.21 (0.071 с.) |
 .
. , де електрична стала
, де електрична стала 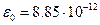 Ф/м, d – відстань між конденсатору, S – площа пластини.
Ф/м, d – відстань між конденсатору, S – площа пластини. .
.