Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Одиночный и групповой образ жизни.Содержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
Одиночный образ жизни характерен для многих видов животных на определенных стадиях жизненного цикла, однако у изолированных особейбыстрее тратятся резервные вещества, не проявляется ряд инстинктов и т.д. Семейный образ жизни усиливает связи между родителями и их потомством, выражается в территориальное поведение животных Эффект группы проявляется как реакция особи на присутствие других особей своего вида Колония – поселения животных, в которых отдельные жизненные функции выполняются сообща, что повышает вероятность выживания особей. Стая – временное объединение животных одного вида, связанное с общностью места обитания или размножения Стадо – группа животных одного вида, обитающая на какой-либо территории или акватории.
Под биоценозом (от лат. биос жизнь, ценоз общий) понимают совокупность живых организмов, населяющих определенную территорию и характеризующихся определенными отношениями между собой и адаптациями к среде обитания. Термин "биоценоз" был предложен немецким гидробиологом К.Мёбиусом в 1877 г. Всю сумму воздействий, которые оказывают друг на друга живые существа, называют биотическими факторами. Непосредственное живое окружение организма составляет его биоценотическую среду. Участок абиотической среды, которую занимает биоценоз, называютбиотопом. Масштабы биоценотических группировок организмов различны - от сообществ лишайников на стволах деревьев или разлагающегося пня до населения целых ландшафтов: лесов, степей, пустынь и т.д. Под экологической структурой биоценоза понимают определенное соотношение экологических групп организмов. Например, соотношение экологических групп организмов по отношению к влажности - гигрофилов (гигрофитов), мезофилов (мезофитов) и ксерофилов (ксерофитов). Биоценозы со сходной экологической структурой могут иметь разный видовой состав, поскольку одни и те же экологические ниши в разных биоценозах могут быть заняты разными видами, сходными по экологии. Такие виды, выполняющие одни и те же функции в сходных типах биоценозов, называютсявикарирующими. Например, одну и ту же экологическую нишу занимают антилопы в саваннах Африки, дикие лошади и куланы в степях Азии. Между организмами в биоценозах возникают разнообразные межвидовые взаимоотношения. Согласно классификации известного отечественного зоолога В.Н.Беклемишева, существует четыре типа биотических взаимоотношений в сообществах. 1. Трофические связи - пищевые связи, при которых один вид питается другим: либо живыми особями, либо их мертвыми остатками, либо продуктами жизнедеятельности. Животные по типу питания делятся на несколько экологических групп: хищники, или зоофаги (питаются животной пищей); сапрофаги (питаются гниющими веществами); некрофаги (питаются трупами животных); копрофаги (питаются экскрементами); фитофаги (питаются растительной пищей). Среди фитофагов выделяют филлофагов (питаются листьями), карпофагов (питаются плодами), ксилофагов (питаются древесиной), ризофагов (питаются корнями). По степени избирательности пищевых объектов выделяют три группы организмов: 1) монофаги - одноядные, 2)олигофаги - ограниченноядные, 3) полифаги - многоядные. 2. Топические связи - взаимоотношения, связанные с местообитанием. Например, конкуренция за место питания, размножения, гнездостроительства. 3. Форические связи - взаимоотношения, связанные с участием одного вида в распространении другого (от слова форезия - перенос одними животными других). Например, рыбы-прилипалы прикрепляются к акулам или черепахам и используют их в качестве "транспорта". Гамазовые клещи часто используют подобным образом различных насекомых. Семена многих растений переносятся различными животными. 4. Фабрические связи - взаимоотношения, при которых один вид использует для своих сооружений другие организмы или их остатки. Птицы используют для строительства гнезд различные растения, пух, шерсть и другой строительный материал биологического происхождения. Согласно другой классификации, между организмами в биоценозах возможны шесть основных типов экологических взаимоотношений (для каждого типа взаимоотношений даются соответствующие обозначения): 1) мутуализм - взаимовыгодные отношения между видами (+; +). Примеры мутуалистических, или симбиотических, взаимоотношений: - лишайники (симбиоз грибов и водорослей; грибы получаютпитательные вещества, водоросли - воду и минеральные вещества); - энтомофильные растения - насекомые-опылители; - кишечные эндосимбионты (бактерии, простейшие) - животные-хозяева (жвачные млекопитающие, термиты); - бобовые растения - клубеньковые бактерии (азот -фиксаторы); - деревья - грибы; 2) комменсализм - одностороннее использование одним видом другого без вреда для последнего (+; 0). Эти взаимоотношения заключаются в том, что деятельность одного вида доставляет пищу или убежище другому. Примеры комменсализма: - рыбы-прилипалы на акулах или черепахах; 3) нейтрализм - независимое существование видов, при котором виды не приносят друг другу ни вреда, ни пользы (0; 0); 4) аменсализм - взаимодействие видов, при котором один не получает ни вреда, ни пользы, а для другого это взаимоотношение отрицательно. Светолюбивые растения, растущие под елью, испытывают угнетение, а для ели это взаимоотношение безразлично (-; 0); 5) конкуренция - форма экологических взаимоотношений, отрицательно сказывающаяся на обоих взаимодействующих видах (-; -). Конкуренция возникает между видами за различные ресурсы (трофические, топические); 6) хищничество и паразитизм - одностороннее использование одним видом (хищник, паразит) другого (хозяин), при котором первый уничтожает или эксплуатирует другого (+; -). Хищничество и паразитизм относятся к пищевым взаимоотношениям. Хищникам для нормального существования необходимы приспособления, позволяющие успешно охотиться на жертв. Поэтомув популяциях хищников естественный отбор будет увеличивать эффективность поиска, ловли и поедания добычи. В то же время в популяции жертв селективное преимущество, как правило, будут иметь те особи, которые успешнее избегают хищников. Долговременная коэволюция (совместная эволюция) хищника и жертвы совершенствует приспособления видов, входящих в эту пару. Сюда относятся как сложные морфологические, так и поведенческие адаптации.
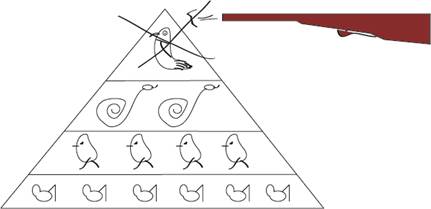
Трофические взаимодействия - это пищевые взаимодействия они регулируют всю энергетику экосистемы в целом. Все организмы делятся на гетеротрофы и автотрофы. Автотрофы – используют неорганические источники (вещества) для своего существования (растения, деревья) участвуют в фотосинтезе (прямая реакция). Гетеротрофы – питаются готовыми органическими веществами (животные, человек) – обратная реакция. ТРОФИЧЕСКАЯ ЦЕПЬ (пищевая цепь, цепь питания), взаимоотношения между организмами, через которые в экосистеме происходит трансформация вещества и энергии; группы особей (бактерии, грибы, растения и животные), связанные друг с другом отношением пища - потребитель. В трофической цепи при переносе потенциальной энергии от звена к звену большая её часть (до 80-90%) теряется в виде теплоты. Поэтому число звеньев (видов) в трофической цепи обычно не превышает 4-5 и, очевидно, чем длиннее трофическая цепь, тем меньше продукция её последнего звена по отношению к продукции начального. В состав пищи каждого вида входит обычно не один, а несколько или много видов, каждый из которых в свою очередь может служить пищей нескольким видам. Поэтому трофические взаимоотношения видов в природе точнее передаются термином трофическая сеть (или паутина). Однако представление о трофической цепи сохраняет своё значение, когда оказывается возможным разнести всех членов сообщества по отдельным звеньям цепи - трофическим уровням. Продуценты (также автотрофные организмы, автотрофы)— организмы, способные синтезировать органические вещества из неорганических. В основном, зелёные растения (синтезируют органические вещества из неорганических в процессе фотосинтеза), однако некоторые виды бактерий-хемотрофов способны на чисто химический синтез органики и без солнечного света. Являются первым звеном пищевой цепи Консументы — организмы, неспособные синтезировать органические вещества из неорганических. Потребляют органические вещества в готовом виде (1-го порядка — растительноядные, 2-го и больших порядков — плотоядные и хищники; всеядные животные). Являются вторым, третьим и далее звеньями пищевой цепи. Редуценты (также деструкторы, сапротрофы, сапрофиты) — организмы, разрушающие остатки мёртвых растений и животных (черви, мокрицы, раки, сомы, грифы) и превращающие их в неорганические соединения (бактерии, грибы). Удаленность организмов от продуцентов одинакова. Они характеризуются определенной формой организации и утилизации энергии. Организмы разных трофических цепей, получающие пищу через равное число звеньев в трофической цепи, находятся на одном трофическом уровне. На каждом трофическом уровне потребленная пища ассимилируется не полностью, т. к. значительная ее часть теряется, тратится на обмен. Поэтому продукция организмов каждого последующего трофического уровня всегда меньше (в среднем в 10 раз) предыдущего. Соотношение различных трофических уровней можно графически изобразить в виде экологической пирамиды. Трофический уровень — это совокупность организмов, занимающих определенное положение в общей цепи питания. К одному трофическому уровню принадлежат организмы, получающие свою энергию от Солнца через одинаковое число ступеней. Так, зеленые растения занимают первый трофический уровень (уровень продуцентов), травоядные животные — второй (уровень первичных консу-ментов), первичные хищники, поедающие травоядных, — третий (уровень вторичных консументов), а вторичные хищники — четвертый (уровень третичных консументов). Трофических уровней может быть и больше, когда учитываются паразиты, живущие на консументах предыдущих уровней. Такая последовательность и соподчиненность связанных в форме трофических уровней групп организмов представляет собой поток вещества и энергии в экосистеме, основу ее организации. Трофическая структура экосистемы. В результате последовательности превращений энергии в пищевых цепях каждое сообщество живых организмов в экосистеме приобретает определенную трофическую структуру. Трофическая структура сообщества отражает соотношение между продуцентами, консументами (отдельно первого, второго и т.д. порядков) и редуцентами, выраженное или количеством особей живых организмов, или пх биомассой, или заключенной в них энергией, рассчитанными на единицу площади в единицу времени. Трофическую структуру обычно изображают в виде экологических пирамид. Эту графическую модель разработал в 1927 г. американский зоолог Чарльз Элтон. Основанием пирамиды служит первый трофический уровень — уровень продуцентов, а следующие этажи пирамиды образованы последующими уровнями — консументами различных порядков. Высота всех блоков одинакова, а длина пропорциональна числу, биомассе или энергии на соответствующем уровне. Различают три способа построения экологических пирамид. 1. Пирамида чисел (численностей) отражает численность отдельных организмов на каждом уровне. Например, чтобы прокормить одного волка, необходимо по крайней мере несколько зайцев, на которых он мог бы охотиться; чтобы прокормить этих зайцев, нужно довольно большое количество разнообразных растений. Иногда пирамиды чисел могут быть обращенными, или перевернутыми. Это касается пищевых цепей леса, когда продуцентами служат деревья, а первичными консументами — насекомые. В этом случае уровень первичных консументов численно богаче уровня продуцентов (на одном дереве кормится большое количество насекомых). 2. Пирамида биомасс — соотношение масс организмов разных трофических уровней. Обычно в наземных биоценозах общая масса продуцентов больше, чем каждого последующего звена. В свою очередь, общая масса консументов первого порядка больше, нежели консументов второго порядка и т.д. Если организмы не слишком различаются по размерам, то на графике обычно получается ступенчатая пирамида с суживающейся верхушкой. Так, для образования 1 кг говядины необходимо 70—90 кг свежей травы. В водных экосистемах можно также получить обращенную, или перевернутую, пирамиду биомасс, когда биомасса продуцентов оказывается меньшей, нежели консументов, а иногда и редуцентов. Например, в океане при довольно высокой продуктивности фитопланктона общая масса в данный момент его может быть меньше, нежели у потребителей-консументов (киты, крупные рыбы, моллюски). Пирамиды чисел и биомасс отражают статику системы, т. е. характеризуют количество или биомассу организмов в определенный промежуток времени. Они не дают полной информации о трофической структуре экосистемы, хотя позволяют решать ряд практических задач, особенно связанных с сохранением устойчивости экосистем. Пирамида чисел позволяет, например, рассчитывать допустимую величину улова рыбы или отстрела животных в охотничий период без последствий для нормального их воспроизведения. 3. Пирамида энергии отражает величину потока энергии, скорость про хождения массы пищи через пищевую цепь. На структуру биоценоза в большей степени оказывает влияние не количество фиксированной энер гии, а скорость продуцирования пищи. Установлено, что максимальная величина энергии, передающейся на следующий трофический уровень, может в некоторых случаях составлять 30 % от предыдущего, и это в лучшем случае. Во многих биоценозах, пищевых цепях величина передаваемой энергии может составлять всего лишь 1 %. В 1942 г. американский эколог Р. Линдеман сформулировал закон пирамиды энергий (закон 10 процентов), согласно которому с одного трофического уровня через пищевые цепи на другой трофический уровень переходит в среднем около 10 % поступившей на предыдущий уровень экологической пирамиды энергии. Остальная часть энергии теряется в виде теплового излучения, на движение и т.д. Организмы в результате процессов обмена теряют в каждом звене пищевой цепи около 90 % всей энергии, которая расходуется на поддержание их жизнедеятельности. Если заяц съел 10 кг растительной массы, то его собственная масса может увеличиться на 1 кг. Лисица или волк, поедая 1 кг зайчатины, увеличивают свою массу уже только на 100 г. У древесных растений эта доля много ниже из-за того, что древесина плохо усваивается организмами. Для трав и морских водорослей эта величина значительно больше, поскольку у них отсутствуют трудноусвояемые ткани. Однако общая закономерность процесса передачи энергии остается: через верхние трофические уровни ее проходит значительно меньше, чем через нижние. Вот почему цепи питания обычно не могут иметь более 3—5 (редко 6) звеньев, а экологические пирамиды не могут состоять из большого количества этажей. К конечному звену пищевой цепи так же, как и к верхнему этажу экологической пирамиды, будет поступать так мало энергии, что ее не хватит в случае увеличения числа организмов. Этому утверждению можно найти объяснение, проследив, куда тратится энергия потребленной пищи (С). Часть ее идет на построение новых клеток, т.е. на прирост (Р). Часть энергии пищи расходуется на обеспечение энергетического обмена 7или на дыхание (i?). Поскольку усвояемость пищи не может быть полной, т.е. 100 %, то часть неусвоенной пищи в виде экскрементов удаляется из организма (F). Балансовое равенство будет выглядеть следующим образом: С = Р + R + F. Учитывая, что энергия, затраченная на дыхание, не передается на следующий трофический уровень и уходит из экосистемы, становится ясным, почему каждый последующий уровень всегда будет меньше предыдущего. Именно поэтому большие хищные животные всегда редки. Поэтому также нет хищников, которые питались бы волками. В таком случае они просто не прокормились бы, поскольку волки немногочисленны. Трофическая структура экосистемы выражается в сложных пищевых связях между составляющими ее видами. Экологические пирамиды чисел, биомассы и энергии, изображенные в виде графических моделей, выражают количественные соотношения разных по способу питания организмов: продуцентов, консументов и редуцентов. Принцип Ле Шателье-Брауна: При внешнем воздействии, выводящем экосистему из состояния устойчивого равновесия, равновесие смещается в том направлении, при котором эффект внешнего воздействия ослабляется.
Вывод: устойчивость экосистемы – это функция биоразнообразия. Следствия: Полностью безотходное производство невозможно, оно равносильно созданию «вечного двигателя». Любая высокоорганизованная биотическая система, которая видоизменяет среду жизни, представляет потенциальную угрозу для более низкоорганизованных систем В биосфере повторное зарождение жизни невозможно – она будет уничтожена существующими организмами Биосфера развивается за счет ресурсов планеты и за счет Солнца. ПРАВИЛО ОДНОГО ПРОЦЕНТА Изменение энергетики природной системы в среднем на 1% выводит последнюю из состояния гомеостаза (равновесия) Переход энергетики процесса за это значение приводит к резким климатическим отклонениям Это может случиться при слишком активной хозяйственной деятельности глобального масштаба и военных конфликтах с применением оружия массового поражения.
Экологическая пирамида - это графическое изображение соотношения различных трофических уровней пищевой цепи. Основание экологической пирамиды составляет уровень продуцентов. Экологические пирамиды могут быть 3 видов:
[ОСТАЛЬНОЕ В ПРЕДЫДУЩЕМ ВОПРОСЕ]
Любой биоценоз динамичен, в нем постоянно происходит изменение в состоянии и жизнедеятельности его членов и соотношении популяций. Все многообразные изменения, происходящие в любом сообществе, можно отнести к двум основным типам: циклические и поступательные. Циклические изменения сообществ отражают суточную, сезонную и многолетнюю периодичность внешних условий и проявления эндогенных ритмов организмов. Суточные преобразования в биоценозах обычно выражены тем сильнее, чем значительнее разница температур, влажности и других факторов среды днем и ночью. Так, в песчаных пустынях Средней Азии жизнь летом в полуденные часы замирает. Даже виды с дневной активностью прячутся от жары в норах, в тени саксаула либо на его ветвях (агамы, ящерки). Ночью пустыня оживает. Ночных и сумеречных животных здесь больше, чем дневных. Многие дневные виды летом переходят к ночному образу жизни (большинство змей, пауки, чернотелки). Активно действуют жуки‑копрофаги, вылетают козодои, домовые сычи, кормятся тушканчики, гекконы, хищничают фаланги, скорпионы, лисицы, корсаки, змеи. Суточные ритмы прослеживаются в сообществах всех зон, от тропиков до тундр. Даже при непрерывном освещении летом в тундре отмечаются суточные ритмы в распускании цветков у растений, питании птиц, в лёте и распределении насекомых и т. п. Сезонная изменчивость биоценозов выражается в изменении не только состояния и активности, но и количественного соотношения отдельных видов в зависимости от циклов их размножения, сезонных миграций, отмирания отдельных генераций в течение года и т. п. На определенное время года многие виды практически полностью выключаются из жизни сообщества, переходя в состояние глубокого покоя (оцепенения, спячки, диапаузы), переживая неблагоприятный период на стадии яиц и семян, перекочевывая или улетая в другие биотопы или географические районы. Сезонной изменчивости подвержена зачастую и ярусная структура биоценоза: отдельные ярусы растений могут полностью исчезать в соответствующие сезоны года, например, травянистый ярус, состоящий из однолетников. Сезонные ритмы сообществ наиболее отчетливо выражены в климатических зонах и областях с контрастными условиями лета и зимы. В слабой форме они, однако, прослеживаются даже в тропических дождевых лесах, где длительность дня, температура и режим влажности очень мало меняются в течение года. Многолетняя изменчивость – нормальное явление в жизни любого биоценоза. Она зависит от изменения по годам метеорологических условий (климатических флюктуации) или других внешних факторов, действующих на сообщество (например, степени разлива рек). Кроме того, многолетняя периодичность может быть связана с особенностями жизненного цикла растений‑эдификаторов, с повторением массовых размножений животных или патогенных для растений микроорганизмов и т. д. Примером могут служить изменения, происходящие на лугах лесостепной Барабы. Количество осадков, выпадающих в Барабинской лесостепи, резко колеблется по годам, и, как правило, ряд засушливых лет чередуется с многолетним периодом обилия осадков. Постепенное понижение уровня почвенно‑грунтовых вод в засушливый период 1950–1955 гг. стимулировало проникновение вслед за ними корневых систем глубокоукореняющихся трав, таких, как колосняк мохнатый, кермек Гмелина и др. У этих видов отсутствовала реакция на иссушение почвы, но при дальнейшем повышении уровня грунтовых вод они пострадали в первую очередь из‑за отмирания затопленных водой корневых окончаний. Засуха особенно сильно сказалась на мезофильных мелкоукореняющихся видах – овсянице красной, полевице тонкой, лапчатке гусиной и других, а также на гидрофитах – ситнике, вейнике и пр. Они или гибли, изреживались, или переходили в состояние покоя. Усиливавшееся в засуху засоление почв послужило причиной массовой гибели от солевого отравления овсяницы красной. Во влажные годы, когда промачивание почвы привело к ее опреснению в верхних слоях, этот вид появился вновь. Высокие температуры и усиление аэрации почвы способствовали развитию бактерий‑нитрификаторов и накоплению нитратов. Это стимулировало массовое размножение в разнотравье осота полевого, одуванчика бессарабского и некоторых других азотолюбивых видов. Когда энергия нитрификации вслед за увеличением влажности почв снизилась, эти виды, не выдержав конкуренции со злаками, перешли в угнетенное состояние. Изменения в фитоценозах были усилены при массовом размножении в 1953 г. нестадных саранчовых. Прямокрылые съели до 80–90 % массы мягколистных злаков: пырея, костра, типчака и др. У жестколистных колосняка, тростника и других они уничтожали всходы по мере их появления. В этот год бурно разрослось непоедаемое саранчовыми разнотравье, для которого была устранена конкуренция со стороны злаков. Последствия размножения саранчовых сказались и на соотношении злаков в последующие два года. В засушливые годы муравьи Lasius niger и Myrmica scabrinodis заселили, кроме возвышенностей, высохшие низинные луга, где начали земляную строительную деятельность, которая повлияла и на условия произрастания растений. Подобные многолетние изменения в составе ценозов имеют тенденцию повторяться вслед за периодическими локальными изменениями климата, которые связаны с изменением общей циркуляции атмосферы, обусловленной, в свою очередь, усилением и ослаблением солнечной активности. Вследствие многостепенной зависимости изменения в сообществах не прямо отражают ритмику космических процессов. Примером обратимых изменений фитоценозов, связанных с особенностями жизненного цикла растений‑эдификаторов, могут быть преобразования в буковом лесу. Взрослые буки с плотно сомкнутыми кронами создают такую густую тень, что в лесу практически отсутствуют подлесок и травяной покров, а сеянцы буков пребывают в угнетенном состоянии и погибают. Когда старые деревья достигают предельного возраста и падают, через появившиеся «окна» на почву проникает солнечный свет и трогаются в рост молодые буки. Некоторое время древостой характеризуется разновозрастностью, пока не выпадут все старые деревья и не выйдут в первый ярус подросшие молодые буки, после чего вновь устанавливается практически одноярусная структура фитоценоза. Весь цикл занимает около двух с половиной столетий. Поступательные изменения в сообществе приводят в конечном счете к смене этого сообщества другим, с иным набором господствующих видов. Причиной подобных смен могут быть внешние по отношению к ценозу факторы, длительное время действующие в одном направлении, например возрастающее в результате мелиорации иссушение болотных почв, увеличивающееся загрязнение водоемов, усиленный выпас скота, вытаптывание лесопарков населением городов и т. п. Возникающие при этом смены одного биоценоза другим называют экзогенетическими. Если при этом усиливающееся влияние фактора приводит к постепенному упрощению структуры сообществ, обеднению их состава, снижению продуктивности, то подобные смены называют дигрессионными или дигрессиями. Так, пастбищные дигрессии на террасовых песках Нижнего Днепра развиваются следующим образом. При умеренном выпасе степь находится на стадии дерновинных злаков. Преобладают типчак, кипец, житняк, ковыль. При дальнейшей усиленной пастьбе возникает стадия стержнекорневых двудольных с господством чаще всего молочая, а также двулетников и однолетников. Дерновинки злаков разбиваются копытами скота, а затем почти совершенно исчезают. На третьей стадии корневищных растений появляются злаки, характерные для сыпучих и слабозаросших песков: песчаный пырей, вейник, осока песчаная. На следующей стадии возникают голые пески с отдельными зарослями псаммофитов, характерных для предыдущего этапа. Таким образом, ковыльно‑типчаковая степь сменяется сыпучими песками. Соответственно меняется и характер животного населения. Эндогенетические смены возникают в результате процессов, происходящих внутри самого сообщества. Закономерный направленный процесс изменения сообществ в результате взаимодействия живых организмов между собой и окружающей их абиотической средой называют сукцессией. Причины возникновения сукцессии. Сукцессия (от лат. successio – преемственность, наследование) – это процесс саморазвития сообществ. В основе сукцессии лежит неполнота биологического круговорота в данном ценозе. Каждый живой организм в результате жизнедеятельности меняет вокруг себя среду, изымая из нее часть веществ и насыщая ее продуктами метаболизма. При более или менее длительном существовании популяций они меняют свое окружение в неблагоприятную сторону и в результате оказываются вытесненными популяциями других видов, для которых вызванные преобразования среды оказываются экологически выгодными. Таким образом, в сообществе происходит смена господствующих видов. Длительное существование биоценоза возможно лишь в том случае, если изменения среды, вызванные деятельностью одних организмов, точно компенсируются деятельностью других, с противоположными экологическими требованиями. В ходе сукцессии на основе конкурентных взаимодействий видов происходит постепенное формирование более устойчивых комбинаций, соответствующих конкретным абиотическим условиям среды. Примерами сукцессий могут быть смены видов при зарастании стоячих водоемов (рис. 153) или барханных песков в пустыне. Процесс сукцессии. Процесс сукцессии, по Ф. Клементсу, состоит из нескольких этапов: 1) возникновения незанятого жизнью участка; 2) миграции на него различных организмов или их зачатков; 3) приживания их на данном участке; 4) конкуренции их между собой и вытеснения отдельных видов; 5) преобразования живыми организмами местообитания, постепенной стабилизации условий и отношений. В настоящее время практически вся доступная жизни поверхность суши занята различными сообществами, и поэтому возникновение свободных от живых существ участков имеет локальный характер. Это или места, освободившиеся в результате отодвигания ледников, отступания уреза воды в водоемах, обвалов, эрозии и т. п., или возникшие в результате деятельности человека, например выноса наверх больших масс глубинных пород при разработке полезных ископаемых. Занос спор, семян растений, проникновение животных на освободившийся участок имеют большей частью случайный характер и зависят от того, какие виды есть в окружающих биотопах. Из числа видов, попавших в новое местообитание, приживаются лишь те, экологическая валентность которых соответствует данному комплексу абиотических условий. Прижившиеся виды постепенно занимают весь новый биотоп, вступая в конкуренцию друг с другом. В результате происходит перестройка видового состава и количественных соотношений разных форм. Параллельно идет процесс преобразования самого местообитания под влиянием развивающегося сообщества. Процесс завершается формированием более или менее стабильной системы с уравновешенным типом биологического круговорота. Сукцессии любого масштаба и ранга характеризуются целым рядом общих закономерностей, многие из которых чрезвычайно важны для практической деятельности человека. В любой сукцессионной серии темпы происходящих изменений постепенно замедляются. Конечным итогом является формирование относительно устойчивой стадии – климаксового сообщества, или климакса. Начальные, пионерные группировки видов отличаются наибольшей динамичностью и неустойчивостью. Климаксовые же экосистемы способны к длительному самоподдерживанию в соответствующем диапазоне условий, так как приобретают такие черты организации биоценозов, которые позволяют поддерживать сбалансированный круговорот веществ. В ходе сукцессии постепенно нарастает видовое многообразие. Это ведет к усложнению связей внутри ценоза, разветвлению цепей питания и усложнению трофической сети, умножению симбиотических отношений, усилению регуляторных возможностей внутри системы. Тем самым уменьшается вероятность слишком сильного размножения отдельных видов и снижается степень доминирования наиболее массовых форм. Чрезвычайное увеличение численности отдельных видов возможно чаще всего на начальных этапах развития сукцессионных серий, когда еще в недостаточной мере сложилась система взаимного регулирования. В незрелых сообществах, т. е. находящихся в начале сукцессионных рядов, преобладают мелкоразмерные виды с короткими жизненными циклами и высоким потенциалом размножения, специализированные на быстром захвате освободившегося пространства. Они обычно обладают широкими расселительными возможностями, позволяющими им первыми проникать на незанятые участки, но малоспособны к конкуренции и длительному удержанию за собой пространства. Постепенно в развивающихся сообществах появляются и закрепляются более крупные формы с длительными и сложными циклами развития. Нарастание экологического разнообразия ведет к более четкому распределению групп организмов по экологическим нишам. В растительном покрове становится сильнее выражена ярусность и мозаичность, создающие основу пространственной структуры наземных экосистем. Усиливается зависимость успешного существования одних видов от биохимических выделений, роста или поведения других: преобладание конкурентных взаимоотношений сменяется преобладанием мутуалистических и трофических зависимостей. В результате сообщества приобретают известную степень автономности и независимости от окружающих условий, не подчиняя свою жизнь флюктуациям внешней среды, а вырабатывая собственные эндогенные ритмы. Не меньшие преобразования происходят и в энергетическом балансе системы (рис. 157). С энергетических позиций сукцессия – такое неустойчивое состояние сообщества, которое характеризуется неравенством двух показателей: общей продуктивности и энергетических трат всей системы на поддержание обмена веществ.
ОРГАНИЗМ И СРЕДА
Аутэкология (факториальная экология) – наука, изучающая взаимоотношения представителей того или иного вида с окружающей средой. Опирается на исследование процессов адаптации видов к окружающей среде, в особенности к факторам парныъ взаимодействия (организм – фактор). Окружающая среда. Понятие «среда» изменяет свою емкость в зависимости от степени конкретизации. СРЕДА: Совокупность внешних условий, в которых совершается какой-либо процесс В применении к истории важное значение среды впервые выдвинуто Монтескье. Позже эта теория обстоятельно разработана Боклем. Шире учение о влиянии природной среды принимается исключительно в психическом или духовно-культурном смысле, как совокупность культурных условий, среди которых живут индивидуумы (Лацарус, Штейнталь), аналогичное понимание среды и у Тэна, тогда как экономический материализм выдвигает на первый план в качестве среды экономичной структуру общества.
ВНЕШНЯЯ СРЕДА. Environment, external envinronment. Точ, что окружает систему и оказывает на нее воздействие. Разграничение системы и среды практически всегда условно, оно
|
||||||||||||||
|
|
Последнее изменение этой страницы: 2017-02-07; просмотров: 794; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.15.195.84 (0.017 с.) |