Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Стимуляция метилирования фосфолипидов.Содержание книги
Поиск на нашем сайте
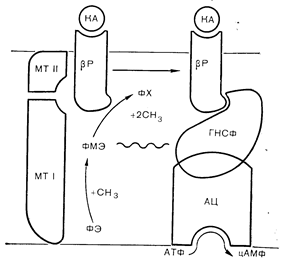
Синтез фосфатидилхолина из фосфатидилэтаноламина в клеточной мембране проходит через два последовательных этапа метилирования, осуществляемых путем переноса метильных групп с S-аденозилметионина под контролем двух ферментов, называемых фосфометилтрансферазами I и II. Первый фермент переносит одну метильную группу, образуя фосфатидилмонометилэтаноламин; как субстрат фосфатидилэтаноламин, так и первый метилтрансферазный фермент локализуются на цитоплазматической стороне клеточной мембраны. Второй фермент переносит еще две метильные группы опять-таки с S-аденозилметионина, образуя фосфатидилхолин. Как продукт фосфатидилхолин, так и вторая метилтрансфераза располагаются на наружной поверхности мембраны. Такое асимметричное распределение ферментов и их субстратов способствует быстрому переносу фосфолипидов через плазматическую мембрану в ходе последовательного метилирования [67]. Внутримембранный синтез промежуточного продукта фосфатидилмонометилэтаноламина вызывает резкие изменения текучести мембраны, создавая условия для ускоренного латерального движения собственных мембранных белков. Стимуляция процесса метилирования фосфолипидов в ретикулоцитах под действием b-адренергических агонистов и снижение вязкости мембраны при этом сопровождается демаскированном скрытых b-адренергических рецепторов, ускорением латерального движения рецепторов и усилением сопряжения между рецепторами и аденилатциклазой. Метилирование мембранных фосфолипидов увеличивает число рецепторов некоторых гормонов, в том числе пролактина и ЛГ. К другим мембранным ферментам и процессам, на которые влияют синтез и транслокация метилированных фосфолипидов, относятся кальцийзависимая АТФаза, хемотаксис лейкоцитов, секреция гистамина тучными клетками и митогенез лимфоцитов (рис. 4—24). В клетках астроцитомы стимуляция b-адренергических и бензодиазепиновых рецепторов повышает метилирование фосфолипидов аддитивным образом, что свидетельствует о независимой стимуляции двумя лигандами метилтрансфераз, локализованных в отдельных доменах вблизи соответствующих рецепторов [68].
Рис. 4 —24. Механизм действия катехоламинов (КА) на метилирование фосфолипидов и мобильность b-адренергических рецепторов (Р). Предполагают, что повышение текучести мембраны облегчает взаимодействие рецепторов с гуанилнуклеотидсопрягающим фактором (ГНСФ) и аденилатциклазой (АЦ) (Hirata, Axelrod [68]). КА — катехоламин: МТ — метилтрансфераза: ФХ — фосфатидилхолин: ФЭ — фосфатидилэтаноламин; ФМЭ — фосфатидилмонометилэтаноламин.
Такие вызываемые лигандами изменения в метилировании фосфолипидов и текучести мембраны, по всей вероятности, отражают общий феномен модификации структуры и функции мембраны при многих формах регуляции клеток гормонами и другими агентами. Они могут объяснять также кратковременное увеличение числа рецепторов гомологичного и других гормонов, наблюдаемое после стимуляции клеток-мишеней. Так, ЛГ в семенниках крысы вызывает временное увеличение рецепторов как самого ЛГ, так и пролактина, а АКТГ вызывает начальное повышение числа рецепторов пролактина в надпочечниках. В каждом из этих случаев увеличение числа рецепторов предшествует изученному процессу потери рецепторов, или «снижающей регуляции», который in vivo начинается через несколько часов после взаимодействия лигандов с рецепторами. Степень, в которой повышение текучести мембраны контролирует рецептор-циклазное сопряжение в клетках-мишенях гормонов, пока не известна, но этот механизм может быть важным компонентом регуляции клеточной чувствительности и реактивности к гормональной стимуляции.
СТЕРОИДНЫЕ ГОРМОНЫ Основные классы стероидных гормонов, секретируемых половыми и надпочечными железами, включают андрогены, эстрогены, прогестины, глюко- и минералокортикоиды. Стадии биосинтеза этих стероидов определяются комплектом ферментов, присутствующих в отдельных стероидогенных клетках-мишенях, а общая скорость синтеза стероидов в каждом случае регулируется одним или несколькими тропными гормонами гипофиза: ЛГ, ФСГ, АКТГ и пролактином.
|
||||
|
|
Последнее изменение этой страницы: 2017-01-26; просмотров: 195; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.138.170.67 (0.008 с.) |