Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
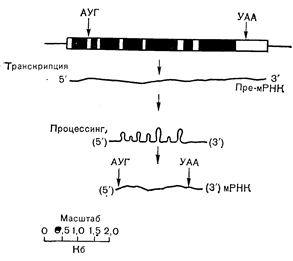
Структура гена и процессинг продуктов транскрипции (мРНК)
Известно, что между транскрипцией РНК на матрице ДНК и появлением транслируемой мРНК в цитоплазме существует несколько стадий. До недавнего времени полагали, что транскрипция приводит к образованию высокомолекулярной РНК, процессинг которой сводится к простому нарезанию специфических молекул мРНК, которые затем и проходят в цитоплазму, где транслируются с образованием соответствующих белков. Однако в настоящее время выяснилось, что стадии процессинга, участвующие в образовании мРНК, более сложны и включают такие модификации, как полиаденилирование 3 -конца и присоединение метилированного гуанозина и к 5 -концу молекулы мРНК. Последовательности-ДНК, определяющие первичную структуру мРНК, также оказались не смежными, как полагали раньше, а, как правило, разъединенными «вставками» (или интронами), которые отсутствуют в комплементарной структуре соответствующих мРНК. Таким образом, мРНК не прямо копируется с генной последовательности, а-транскрибируется вначале в виде гораздо более крупного продукта, из которого в ходе процессинга вырезаются промежуточные последовательности с образованием зрелой мРНК, непосредственно транслирующейся на рибосомном уровне [104]. Присутствие промежуточных последовательностей ДНК в эукариотических генах впервые было обнаружено в генах b-глобулина, легких цепей иммуноглобулинов и альбумина. Позднее такие вставки были найдены также в генах овальбумина, кональбумина и овомукоида, равно как и в других генах специфических белков [1, 104]. Современная точка зрения на организацию овальбуминового гена, отраженная на рис. 4—28, заключается в том, что структурный ген разъединяется по меньшей мере 7 промежуточными последовательностями. Стимуляция эстрогеном вызывает координированное повышение транскрипции структурных и промежуточных последовательностей овальбуминового гена с накоплением как высокомолекулярной РНК, содержащей ов альбуминовую последовательность, так и цитоплазматической овальбуминовой мРНК [105]. Молекулы крупного предшественника мРНК, содержащего промежуточные последовательности, подвергаются воздействию ферментов с вырезанием вставок и последующим сшиванием, что приводит к образованию зрелой мРНК (см. рис. 4—28). Функция промежуточных последовательностей (или интронов) в эукариотических структурных генах не выяснена. Одно из предположений заключается в том, что они ограничивают участки гена, кодирующие домены белка, ответственные за отдельные части его общей функции, и что структурные участки ДНК (или экзоны) сближались в процессе эволюции при участии интронов [106]. Применимы ли эти объяснения к овальбумину, не ясно, поскольку специфические функции этого белка яйцеводов не известны. Однако данные о том, что ген кональбумина (овотрансферрина) содержит 17 экзонов, позволяют проверить эту концепцию путем анализа функциональных доменов, принимающих участие в секреции (сигнальная последовательность) белка и связывании железа [107].
Рис. 4—28. Структурная организация и транскрипция гена овальбумина. Эстроген стимулирует экспрессию всего гена, а промежуточные последовательности затем вырезаются со сшиванием отрезков и образованием зрелой мРНК овальбумина цыплят (Chan и соавт. [1] в модификации). ов — овальбумин; светлый участок — структурный ген темный участок — вставочные последовательности ДНК: линия — боковые последовательности ДНК.
Анализ генома клеток яйцеводов вблизи овальбуминового гена позволил недавно обнаружить два соседних гена с последовательностями, гомологичными овальбуминовому гену [108]. Эти два дополнительных гена имели и интроны, сходные с таковыми в гене овальбумина, и экспрессировались при стимуляции эстрогенами, хотя и в меньшей степени, чем овальбуминовый ген. Такой кластер родственных генов мог бы возникнуть в процессе эволюции-путем параллельной сборки из смеси родственных и неродственных участков или путем удвоения исходного гена, в котором сочетание нитронов и экзонов было зафиксировано с самого начала [109]. Отражает ли разная экспрессия трех генов при действии эстрогенов различия в эффективности транскрипции или различия в последующем процессинге и стабилизации мРНК, пока не установлено. Организация овальбуминовой области генома аналогична таковой генов глобина, и кластеризация структурно и функционально близких генов может быть общей чертой генома эукариотических клеток [108].
ТИРЕОИДНЫЕ ГОРМОНЫ Тиреоидные гормоны оказывают многочисленные и разнообразные эффекты на дифференцировку, развитие и метаболический гомеостаз, контролируя синтез и активность регуляторных белков, в том числе ключевых ферментов метаболизма, гормонов и рецепторов. Известное действие тиреоидных гормонов на потребление кислорода определяется отчасти стимуляцией натриевого насоса за счет индукции мембранного фермента: натрий-, калийзависимой АТФазы [110а]. Этот и другие метаболические эффекты тиреоидных гормонов зависят от гормональной индукции синтеза РНК, осуществляемой путем регуляции экспрессии генов на ядерном уровне. Хотя постулировались прямые влияния тиреоидных гормонов и на клеточную мембрану и на митохондрии, но многие из эффектов этих гормонов на процессы метаболизма опосредуются, по-видимому, активацией ядерных реакций связывания с увеличением образования специфических РНК. Основным йодтиронином, секретируемым щитовидной железой, является тироксин (Т4), которому сопутствует небольшое количество активного трийодтиронина (Т3). В тканях-мишенях Т4 дейодируется в Т3, который представляет собой главную внутриклеточную форму гормона. Это превращение происходит на плазматической мембране и в эндоплазматическом ретикулуме, и в разных тканях варьирует по степени, будучи интенсивным в передней доле гипофиза и менее выраженным в печени и почках. Большая зависимость гипофиза от превращения циркулирующего в крови Т4 в Т3 могла бы определяться потребностью в быстром увеличении гипофизарной продукции ТТГ при небольших изменениях в секреции Т4 щитовидной железой. Большая часть циркулирующих в крови тиреоидных гормонов связана с белками плазмы: тироксинсвязывающим глобулином (ТСГ), тироксинсвязывающим преальбумином (ТСПА) и альбумином. Все они связывают Т4 более прочно, чем Т3, причем ТСГ переносит около 70% связанного Т4 плазмы; из остального количества около 10% связано с ТСПА, а 20% с альбумином [111]. Как и в отношении стероидных гормонов, именно свободные тиреоидные гормоны ответственны за контроль активности клеток-мишеней, и функция связывающих белков, помимо того, что они являются резервуаром циркулирующего гормона, неясна. Интересной особенностью ТСПА является то, что он в некоторых отношениях напоминает клеточный рецептор тиреоидного гормона, в том числе присутствием предположительного ДНК-связывающего участка, богатого заряженными аминокислотами и триптофановыми остатками.
Рис. 4—29. Эффекты и взаимопревращение Т4 и Т3 В клетках, реагирующих на поступление тиреоидных гормонов. Процесс активации ядра зависит от связывания Т3 с ядерными рецепторами, а не от отдельно существующих цитоплазматических рецепторов, как это имеет место в клетках-мишенях стероидных гормонов (Eberhardt и соавт. [111] в модификации).
|
||||||
|
|
Последнее изменение этой страницы: 2017-01-26; просмотров: 164; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.222.69.152 (0.007 с.) |