Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Синтез эпифизарных продуктов
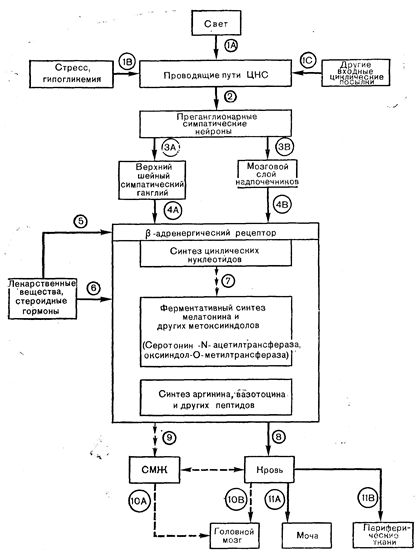
Иннервация эпифиза обладает некоторыми уникальными особенностями. Она почти целиком представлена автономными постганглионарными волокнами, берущими начало в верхнем шейном ганглии. Эти волокна образуют синапсы на паренхиматозных клетках эпифиза. Считается, что афферентные волокна к верхнему шейному ганглию берут начало либо из нижнего акцессорного зрительного тракта, либо из системы ретиногипоталамических связей. В симпатических нервных окончаниях эпифиза присутствует норадреналин, а, кроме того, как в этих окончаниях, так и в пинеалоцитах, обнаружен серотонин. Он служит предшественником одного из эпифизарных «гормонов» мелатонина. Пинеалоциты захватывают триптофан, превращая его в 5-окситриптофан и затем в серотонин. Превращение серотонина в мелатонин требует двух ферментативных этапов; на первом из них участвует серотонин-N-ацетилтрансфераза, которая превращает серотонин в N-ацетил-серотонин, а на втором — оксииндол-О-метилтрансфераза (ОИОМТ), которая переносит метильную группу на N-ацетилсеротонин, образуя мелатонин. Активность ОИОМТ сконцентрирована в основном в эпифизе, хотя ее присутствие обнаружено и в других органах, таких, как сетчатка и гардерианова железа — небольшое образование в глазнице человека. Концентрация серотонина, мелатонина, серотонин-N-ацетилтрансферазы и ОИОМТ в эпифизе подвержена циркадным колебаниям. У крыс максимальный уровень серотонина и обоих ферментов достигается в светлое дневное время и резко снижается вскоре после наступления темноты, когда концентрация мелатонина, низкая в светлое время суток, начинает повышаться [61]. Связь освещения, симпатической нервной системы и эпифизарных индолов и гормонов представлена на рис. 6—9 [62]. Отсутствие света (темнота) повышает постганглионарную симпатическую активность и тем самым высвобождает норадреналин из окончаний на эпифизарных клетках. Возникающая при этом активация аденилатциклазы пинеалоцитов приводит к повышению активности триптофангидроксилазы (превращающей триптофан в 5-окситриптофан), серотонин-М-ацетилтрансферазы и ОИОМТ, что обусловливает снижение концентрации серотонина и увеличение концентрации мелатонина. При воздействии света наблюдается противоположный эффект. Как показано на рисунке, другие воздействия, активирующие симпатическую нервную систему, могут преодолевать ингибирующее влияние освещения. Инсулиновая гипогликемия или иммобилизационный стресс приводят к повышению активности ферментов эпифиза.
Циркадные колебания концентрации мелатонина в плазме и моче отражают колебания его содержания в эпифизе. Интересно, что как у крыс, которые являются ночными животными, так и у человека, организм которого активен преимущественно днем, максимального значения концентрация мелатонина достигает в темное время суток. У эпифизэктомированных крыс концентрация мелатонина в плазме и моче резко снижается, но все еще поддается определению; при этом суточные колебания этих концентраций исчезают. Уровень мелатонина может зависеть и от фазы менструального цикла, будучи наибольшим в период менструаций и наименьшим в период максимального выброса ЛГ. Описаны и другие эпифизарные гормоны, кроме мелатонина-К ним относятся другие метоксииндолы, аргинин-вазотоцин и, возможно, ГнРГ, ТРГ и ренин. Следует отметить, что синтез этих соединений в эпифизе еще не доказан. Вполне может быть, что существуют и дополнительные эпифизарные гормоны, определяющие некоторые непонятные эффекты эпифизэктомии и экстрактов эпифиза на эндокринную функцию. Эпифизэктомия у экспериментальных животных приводит к появлению признаков стимуляции системы гипофиз—половые железы как у самок, так и у самцов, Рис. 6—9. Диаграмма факторов, контролирующих секрецию эпифизарного гормона и поступление его (или их) к органам-мишеням. Главным входным фактором, контролирующим синтез мелатонина в эпифизе, является норадреналин, высвобождаемый постганглионарными симпатическими нервами (3А, 4А). Высвобождение норадреналина тормозится в условиях освещения (влияние света передается через сетчатку и акцессорный зрительный тракт) и может увеличиваться при стрессе (1В), вызывающем генерализованную активацию симпатоадреналовой системы; последнее влияет на эпифиз и через высвобождаемый в кровь адреналин (3В, 4В). В отсутствие циклических изменений освещенности окружающей среды (например, при длительном содержании животных в темноте) синтез и секреция мелатонина сохраняют циркадную ритмичность, что требует интактности симпатических нервов эпифиза (3А, 4А): следовательно, состояние симпатических нервов может определяться и другими входными циклическими посылками (1С), имеющими, вероятно, эндогенное происхождение (т. е. возникающими в головном мозге). Высвобождаемые норадреналин и адреналин действуют черезадренергические рецепторы на поверхности эпифизарных клеток (4А, 4В), активируя аденилатциклазу и повышая тем самым уровень цАМФ в эпифизе; это вещество может опосредовать симпатический нервный контроль ферментов, катализирующих биосинтез мелатонина. Препараты, прямо или опосредованно активирующие рецепторы норадреналина (например, L-ДОФА),.также могут ускорять биосинтез мелатонина. Кроме того, его продукция может изменяться под влиянием эстрогенов и близких стероидных гормонов, механизм действия которых не до конца ясен.
Считается, что кроме мелатонина гормональной активностью обладают еще две группы эпифизарных продуктов: другие метоксииндолы, такие, как 5-метокситриптофол, и пептиды, такие, как 8-аргининвазотоцин. О регуляции синтеза пептидов эпифиза ничего не известно. У человека и теленка концентрация мелатонина в спинномозговой жидкости (СМЖ) выше, чем одновременное содержание его в крови; следовательно, гормон может секретироваться прямо в СМЖ, а не в кровоток. Главный объект его действия расположен в головном мозге (10А. 10В), но он может непосредственно влиять также на гипофиз и другие периферические органы (11В). У всех до сих пор изученных видов уровни мелатонина в СМЖ. крови и моче обнаруживают параллельные суточные колебания с максимумом в темные часы. ("Wurtinan R. «Г., Mosko-witz M. A., The pineal organ. — N. Engl. J. Med., 1977, 296, 1329).
Эпифизэктомия в препубертатном возрасте приводит к преждевременному половому созреванию, а в гипофизе эпифизэктомированных животных обнаружено повышение количества ЛГ и ФСГ. У таких животных отмечалось и увеличение массы щитовидной железы, захвата 131I и секреции тиреоидных гормонов, а также появление признаков активации коры надпочечников, что проявлялось увеличением их массы и ускорением секреции гормонов (альдостерона, кортикостерона). Введение мелатонина купирует многие, но не все эти эффекты. Острое внутрижелудочковое введение мелатонина крысам снижает концентрацию кортикостерона в крови, а инъекция вазотоцина в III желудочек мозга кошек тормозит выделение кортикотропин-рилизинг фактора, предположительно, серотонинергическиу механизмом. Место приложения действия мелатонина, определяющее его эффекты, изучалось в основном в половой сфере. Мелатонин тормозит реакцию гипофизарного ЛГ на ГнРГ как in vivo, так и in vitro. Считают, что он подавляет функцию половых желез, действуя на ГнРГ в ЦНС. Показано, что внутрибрюшинное введение мелатонина повышает концентрацию серотонина в мозге, причем установлено также поглощение мелатонина ЦНС. Неясно, является ли это физиологическим или фармакологическим эффектом. К другим доказательствам влияния мелатонина на ЦНС относятся данные о том, что у кошек он вызывает характерные для определенных стадий сна сдвиги на ЭЭГ, причем сообщалось, что у кошек и человека он оказывает седативное действие и иногда даже вызывает сон. Мелатонин обнаружен в СМЖ, где его концентрация в равнозначных условиях превышает таковую в периферической крови. Это указывает (хотя и не доказывает) на возможность секреции мелатонина эпифизом непосредственно в СМЖ. В определенных условиях у животных некоторых видов наблюдали и стимулирующее половые железы действие мелатонина. Роль мелатонина в регуляции функции половых желез у человека изучена недостаточно. Не следует считать, будто все эффекты эпифизэктомии опосредуются выпадением секреции мелатонина, поскольку, как уже отмечалось, в эпифизе присутствуют другие вещества, которые могут обладать свойствами гормонов.
|
||||||
|
|
Последнее изменение этой страницы: 2017-01-26; просмотров: 131; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.116.36.192 (0.025 с.) |