Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Палеонтологические доказательства эволюцииСодержание книги
Похожие статьи вашей тематики
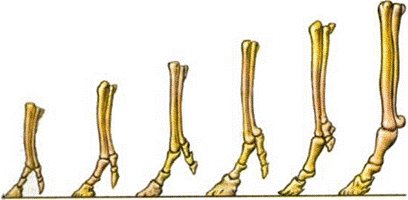
Поиск на нашем сайте Палеонтология ― наука об ископаемых животных и растениях. Развитие хордовых осуществлялось поэтапно. Вначале возникли низшие хордовые, затем последовательно во времени возникли рыбы, амфибии, рептилии. Рептилии, в свою очередь, дали начало млекопитающим и птицам. На заре своего эволюционного развития млекопитающие были представлены небольшим числом видов, в то время процветали рептилии. Позднее резко увеличилось число видов млекопитающих и птиц, а большинство видов рептилий исчезло. Таким образом, палеонтологические данные указывают на смену форм животных. В отдельных случаях палеонтология указывает на причины эволюционных преобразований. В этом отношении интересна эволюция лошадей. Современные лошади произошли от мелких всеядных предков, живших 60-70 млн. лет назад в лесах и имевших пятипалую конечность. Изменение климата на Земле, повлекшее за собой сокращение площадей лесов и увеличение размеров степей, привело к тому, что предки современных лошадей начали осваивать новую среду обитания ― степи. Необходимость защиты от хищников и передвижений на большие расстояния в поисках хороших пастбищ привела к преобразованию конечностей ― уменьшению числа фаланг вплоть до одной. Параллельно изменению конечностей происходило преобразование всего организма: увеличение размеров тела, изменение формы черепа и усложнение строения зубов, возникновение свойственного травоядным млекопитающим пищеварительного тракта и многое другое. В результате изменения внешних условий под влиянием естественного отбора произошло постепенное превращение мелких пятипалых всеядных животных в крупных травоядных. Богатейший палеонтологический материал ― одно из наиболее убедительных доказательств эволюционного процесса, длящегося на нашей планете уже более 3 млрд. лет. Рис. 56. Исторический ряд изменений в строении передней конечности лошади
Биогеографическое доказательство эволюции Ярким свидетельством произошедших и происходящих эволюционных изменений является распространение животных и растений по поверхности нашей планеты. Еще в эпоху великих географических открытий путешественников и натуралистов поражало разнообразие животных в дальних странах, особенности их распространения. Однако лишь Уоллесу удалось привести все сведения в систему и выделить шесть биогеографических областей: Палеоарктическую, Неоарктическую, Индо-Малайскую, Эфиопскую, Неотропическую, Австралийскую.
Рис. 57. Карта биогеографических зон
Сравнение животного и растительного мира разных зон дает богатейший научный материал для доказательства эволюционного процесса. Фауна и флора Палеоарктической (Евроазиатской) и Неоарктической (Североамериканской) областей, например, имеют много общего. Это объясняется тем, что в прошлом между названными областями существовал сухопутный мост ― Берингов перешеек. Неоарктическая и Неотропическая области, напротив, имеют мало общих черт, хотя в настоящее время соединены Панамским перешейком. Это объясняется изолированностью Южной Америки в течение нескольких десятков миллионов лет. После возникновения Панамского моста лишь немногим южноамериканским видам удалось проникнуть на север (дикобраз, броненосец, опоссум). Североамериканские виды преуспели в освоении южноамериканской области несколько больше. Ламы, олени, лисы, выдры, медведи проникли в Южную Америку, но не оказали существенного влияния на ее уникальный видовой состав. Интересен и своеобразен животный мир Австралийской области. Известно, Австралия обособилась от Южной Азии еще до возникновения высших млекопитающих. Таким образом, распределение видов животных и растений по поверхности планеты и их группировка в биогеографические зоны отражает процесс исторического развития Земли и эволюции живого. Островные фауна и флора Для понимания эволюционного процесса интерес представляют фауна и флора островов. Состав их фауны и флоры полностью зависит от происхождения островов. Острова могут быть материкового происхождения, т.е. представлять собой результат обособления части материка или океанического происхождения (вулканические или коралловые). Материковые острова характеризуются фауной и флорой, близкой по составу к материковой. Однако, чем древнее остров и чем более значительна водная преграда, тем больше обнаруживается отличий. Британские острова отделились от Европы совсем недавно и имеют фауну и флору, идентичную европейской. На давно обособившихся островах процесс расхождения видов значительно заметнее. На Мадагаскаре, например, нет типичных для Африки крупных копытных: быков, антилоп, носорогов, зебр. Нет и крупных хищников (львов, леопардов, гиен), высших обезьян (павианов, мартышек). Однако много низших приматов ― лемуров, которые нигде больше не встречаются. Совершенно иная картина обнаруживается при рассмотрении фаун океанических островов. Их видовой состав очень беден. На большинстве таких островов отсутствуют наземные млекопитающие и амфибии, неспособные преодолеть значительные водные преграды. Вся фауна океанических островов ― результат случайного занесения на них некоторых видов, обычно птиц, рептилий, насекомых. Представители таких видов, попавшие на океанические острова, получают широкие возможности для размножения. Например, на Галапагосских островах из 108 видов птиц 82 эндемичны (т.е. нигде больше не встречаются) и все 8 видов рептилий характерны только для этих островов. На Гавайских островах обнаружено большое разнообразие улиток, их 300 эндемичных видов принадлежат одному роду. Огромное количество разнообразных биогеографических факторов указывают на то, что особенности распределения живых существ на планете тесно связаны с преобразованием земной коры и с эволюционными изменениями видов. Молекулярные доказательства эволюции В настоящее время практически завершена полная расшифровка генома (совокупности всех генов) человека и геномов ряда животных, растений и микроорганизмов. Известна полная последовательность нуклеотидов в ДНК у огромного числа видов живых организмов. Сравнение этих последовательностей дает новый ключ к построению родословной жизни на Земле. Многие мутации представляют собой замены одних нуклеотидов на другие. Мутации возникают, как правило, во время репликации ДНК. Отсюда следует, что, чем больше поколений появилось на свет со времени дивергенции двух видов от общего предка, тем больше случайных замен нуклеотидов должно было накопиться в геномах этих дочерних видов. Общий предок человека и мыши существовал более 80 миллионов лет назад. Когда мы сравниваем нуклеотидные последовательности генов (например, гена бета-глобина), мы видим, что различий между генами человека и шимпанзе гораздо меньше, чем между генами человека и мыши. Количественная оценка этих различий позволяет построить генеалогическое древо, показывающее родство различных таксонов (видов, отрядов, семейств, классов), и определить относительное время их дивергенции. В основном это древо совпадает с теми, что были построены на основе морфологических, эмбриологических и палеонтологических данных. Однако в некоторых случаях обнаруживаются поразительные вещи. Оказалось, что киты и парнокопытные гораздо более близкие родственники, чем парнокопытные и непарнокопытные. Африканский златокрот филогенетически ближе к слону, чем к нашим кротам. Современные методы молекулярной генетики позволяют анализировать гены не только ныне живущих организмов, но и давно вымерших видов, используя следы ДНК в ископаемых останках. Это помогает проследить пути эволюции жизни на Земле. Синтетическая теория эволюции (СТЭ) — современная эволюционная теория, которая является синтезом различных дисциплин, прежде всего, генетики и дарвинизма. СТЭ также опирается на палеонтологию, систематику, молекулярную биологию и другие. В 1930-40-е годы быстро произошел широкий синтез генетики и дарвинизма. Генетические идеи проникли в систематику, палеонтологию, эмбриологию, биогеографию. Термин «современный» или «эволюционный синтез» происходит из названия книги Дж. Хаксли «Evolution: The Modern synthesis» (1942). Выражение «синтетическая теория эволюции» в точном приложении к данной теории впервые было использовано Дж. Симпсоном в 1949 году. Авторы синтетической теории расходились во мнениях по ряду фундаментальных проблем и работали в разных областях биологии, но они были практически единодушны в трактовке следующих основных положений: · элементарной единицей эволюции считается локальная популяция; · материалом для эволюции являются мутационная и рекомбинационная изменчивость; · естественный отбор рассматривается как главная причина развития адаптаций, видообразования и происхождения надвидовых таксонов; · дрейф генов и принцип основателя выступают причинами формирования нейтральных признаков; · вид есть система популяций, репродуктивно изолированных от популяций других видов, и каждый вид экологически обособлен; · видообразование заключается в возникновении генетических изолирующих механизмов и осуществляется преимущественно в условиях географической изоляции. Таким образом, синтетическую теорию эволюции можно охарактеризовать как теорию органической эволюции путем естественного отбора признаков, детерминированных генетически. Активность американских создателей СТЭ была столь высока, что они быстро создали международное общество по изучению эволюции, которое в 1946 стало учредителем журнала «Evolution». Журнал «American Naturalist» вновь вернулся к публикации работ по эволюционной тематике, делая акцент на синтезе генетики, экспериментальной и полевой биологии. В результате многочисленных и самых разнообразных исследований основные положения СТЭ прошли не только успешную проверку, но и видоизменялись, дополнялись новыми идеями. В 1942 немецко-американский орнитолог и зоогеограф Э. Майр издал книгу «Систематика и происхождение видов», в которой была последовательно развита концепция политипического вида и генетико-географическая модель видообразования. Майр предложил принцип основателя, который в окончательной форме был им сформулирован в 1954. Если дрейф генов, как правило, дает причинное объяснение формированию нейтральных признаков во временном измерении, то принцип основателя в пространственном. После публикации трудов Добржанского и Майра систематики получили генетическое объяснение тому, во что они уже давно верили: подвиды и близкородственные виды различаются по адаптивно-нейтральным признакам. Ни один из трудов по СТЭ не может сравниться с упомянутой книгой английского экспериментального биолога и натуралиста Дж. Хаксли «Evolution: The Modern synthesis» (1942 год). Труд Хаксли по объему анализируемого материала и широте проблематики превосходит даже книгу самого Дарвина. Хаксли на протяжении многих лет держал в уме все направления в развитии эволюционной мысли, внимательно следил за развитием родственных наук и имел личный опыт генетика-экспериментатора. Видный историк биологии Провин так оценил труд Хаксли: «Эволюция. Современный синтез» была наиболее всесторонней по теме и документам, чем другие работы на эту тему. Книги Холдейна и Добржанского были написаны главным образом для генетиков, Майра для систематиков и Симпсона для палеонтологов. Книга Хаксли стала доминантной силой в эволюционном синтезе". По объёму книга Хаксли не имела себе равных (645 страниц). Но самое интересное состоит в том, что все основные идеи, изложенные в книге, были очень ясно выписаны Хаксли на 20 страницах ещё в 1936, когда он послал адрес Британской ассоциации содействия науки под названием «Natural selection and evolutionary progress». В этом аспекте ни одна из публикаций по эволюционной теории, вышедшая в 1930-40-х годах, не может сравниться со статьей Хаксли. Хорошо чувствуя дух времени, Хаксли писал: «В настоящее время биология находится в фазе синтеза. До этого времени новые дисциплины работали в изоляции. Сейчас проявилась тенденция к унификации, которая является более плодотворной, чем старые односторонние взгляды на эволюцию» (1936). Ещё в трудах 1920-х годов Хаксли показал, что наследование приобретенных признаков невозможно; естественный отбор действует как фактор эволюции и как фактор стабилизации популяций и видов (эволюционный стазис); естественный отбор действует на малые и крупные мутации; географическая изоляция — важнейшее условие видообразования. Кажущаяся цель в эволюции объясняется мутациями и естественным отбором.
Практическая работа 11 Цель работы Обучаемый должен знать историю становления эволюционных идей о развитии органического мира Земли. Обучаемый должен уметь использовать доказательства эволюции и положения современной синтетической теории в обосновании исторического развития органического мира на Земли.
Бюджет времени На изучение темы отводится 6 часов. Из них 2 часа – лекции, 2 часа практические занятия и 2 часа – на самоподготовку.
ЛИТЕРАТУРА - Пехов А.П. Биология с основами экологии. - СПб.: Издательство «Лань», 2000. - 672 с. - Биология. В2 кн./ Под ред. В.Н.Ярыгина.- М.: Высш. Шк., 2001. Темы рефератов 1. Карл Линней и его школа. 2. Ж.Б. Ламарк – автор первой целостной концепции эволюции. 3. Ч. Дарвин. Вопросы для самоконтроля 1. Ранние этапы развития эволюционных представлений. 2. Теория эволюции Ж.Б. Ламарка. 3. Теория эволюции Ч. Дарвина. 4. Современная синтетическая теория эволюции.
Тема 12 УЧЕНИЕ О МИКРОЭВОЛЮЦИИ И ВИДООБРАЗОВАНИИ
В центре представлений Ч. Дарвина об эволюции органического мира стоит вид. Вид объединяет организмы, сходные по морфологическим признакам. Это позволяет отличить одну группу организмов от другой. Такое понимание вида помогло классифицировать огромное разнообразие описанных форм организмов и сыграло положительную роль в развитии биологии. Накопление к концу XVII в. сведений о многообразии форм животных и растений привело к представлению о видах как о вполне реальных группах особей. Основополагающая работа о видах была проведена К. Линнеем, который заложил основы систематики. До Ч. Дарвина большинство биологов считали, что виды неизменны с момента создания их Творцом. Эта "неизменность" являлась решающим аргументом всех антиэволюционных учений. По мере углубления и расширения исследований различных видов ученые перестали ограничиваться редкими музейными экспонатами, а использовали более обширный ареал. Было установлено, что вид включает организмы, характеризующиеся высокой степенью изменчивости различных признаков. В результате этого некоторые ранее самостоятельные виды были объединены в один общий вид. Некоторые ученые даже усомнились в реальности существования вида. Они высказывали мнение, что в действительности видов как реальных биологических единиц в природе не существует, и что границы между видами ученые устанавливают более или менее произвольно. Но все дальнейшие исследования доказали, что виды реально существуют, они являются важнейшей ступенью организации живой природы. Реальность вида в первую очередь доказывается системой критериев, которые позволяют достаточно четко отличить один вид от другого.
|
||
|
|
Последнее изменение этой страницы: 2016-12-10; просмотров: 1923; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.108 (0.009 с.) |