Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Электрогенез в синоатриальном узлеСодержание книги
Поиск на нашем сайте
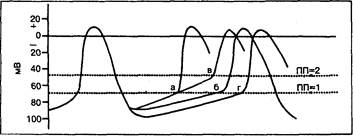
Спонтанная деполяризация (фаза 4) является той электрофизиологической характеристикой, которая отличает пейсмекерные клетки от всех других клеток организма. Синусовый узел способен быть доминирующим водителем сердечного ритма благодаря двум основным электрофизиологическим особенностям его клеток: 1) низкий уровень мембранного потенциала покоя или максимального диастолического потенциала (—60 мВ); 2) высокая скорость нарастания деполяризации в фазу 4. Оценка ионных изменений, обусловливающих повышение спонтанной диастолической деполяризации в синусовом узле, до недавнего времени была затруднена из-за отсутствия метода фиксации потенциала в узловой ткани [9]. В результате интенсивных микроэлектродных исследований, однако, в качестве возможных механизмов, ответственных за фазу 4 деполяризации, были предложены следующие изменения мембранных характеристик (одной из них или в сочетании): 1) снижение проницаемости для выходящего калия; 2) повышение проницаемости для входящего натрия; 3) снижение активности натриевого насоса; 4) повышение проницаемости для входящего кальция. Хотя наиболее распространенным объяснением возникновения спонтанной пейсмекерной деполяризации является наличие потенциалов, превышающих зависимое от времени уменьшение выходящего калиевого тока, ряд фактов свидетельствует о неприменимости этой теории к синусовому узлу. Во-первых, деполяризация в фазу 4 клеток синусового узла происходит в том диапазоне потенциалов, где пейсмекерный ток полностью активируется в клетках-пейсмекерах, зависимость которых от уменьшения выходящего калиевого тока доказана (т. е. в волокнах Пуркинье) [10—12]. Во-вторых, наклон деполяризации в фазу 4 в клетках синусового узла ("по сравнению с волокнами Пуркинье) относительно мало изменяется под угнетающим влиянием повышения внеклеточной концентрации калия [13]. В настоящее время получены убедительные данные о том, что пассивный натриевый ток в лучшем случае играет лишь минимальную роль в инициации возбуждения синусового узла. Характерно, что изменения внеклеточной концентрации натрия практически не влияют на наклон деполяризации в фазу 4 [14]. Активный транспорт натрия, по-видимому, также мало способствует генерированию импульса в синусовом узле; ни тетродотоксин, ни замена ионов натрия на литий, блокирующие работу натриевого насоса, не оказывают значительного влияния на наклон деполяризации в фазу 4 [15—17]. Понимание механизмов генерирования импульса в синусовом узле было существенно расширено в последнее время благодаря осознанию важной роли медленных каналов [18, 19]. В генерировании медленного входящего тока вскоре после первоначального быстрого входящего тока в клетках синусового и атриовентрикулярного узлов, как полагают, принимают участие и ионы натрия, и ионы кальция. Хотя порог активации медленного тока (от —40 до —30 мВ) является положительным для диапазона потенциалов, в пределах которого в основном возникает пейсмекерность, результаты микроэлектродных исследований говорят о том, что медленный ток может играть важную роль в генерировании импульса в синусовом узле. Характерно, что ингибиторы медленных каналов, такие, как D-600, Mg2+ и верапамил, угнетают деполяризацию в фазу 4 в клетках синусового узла [20—24]. Выдвинуто предположение, что в ходе диастолической деполяризации происходит очень медленная инактивация медленного входящего тока, активированного во время фазы плато предыдущего потенциала действия, что объясняет сохранение этого тока при максимальном диастолическом мембранном потенциале синусового узла [25]. Современные теории и математические модели, предлагаемые для объяснения исключительно сложной связи между различными ионами и токами, участвующими в развитии медленной диастолической деполяризации в пейсмекерных клетках синусового узла, должны учитывать данные последних исследований, касающихся роли анионных токов. Входящий ток ионов хлора, по-видимому, действительно принимает участие в медленной диастолической деполяризации клеток синусового узла [26—28]. Анионная проницаемость мембран в синусовом узле значительно выше, чем в волокнах Пуркинье. Замена анионов хлора во внеклеточной среде другими, более легко проникающими через мембрану анионами (например, брома) приводит к ускорению спонтанной диастолической деполяризации в изолированных клетках синусового узла. Замена анионов хлора хуже проникающими через мембрану анионами (например, анионами метилсульфата) замедляет спонтанную диастолическую деполяризацию. Хотя вклад ионов хлора в фазу 4 деполяризации в каждый данный момент ее развития остается неясным, он, вероятнее всего, определяет лишь часть тока, ответственного за медленную диастолическую деполяризацию клеток синусового узла. Как показывают исследования методом фиксации потенциала, точное определение того или иного объема ионных токов, участвующих в генерировании диастолической деполяризации в синусовом узле, затруднено также тем, что клетки в разных частях узла имеют различные электрофизиологические характеристики, изменяющиеся в зависимости от участия конкретного вида ионов. «Зона доминирующего водителя ритма», по-видимому, расположена в центре узла, где группа приблизительно из 5000 клеток с идентичной синхронной активностью, максимальный диастолический потенциал которых составляет около —50 мВ, имеет короткие периоды полупути между максимальным диастолическим потенциалом и пиком потенциала действия. По чисто техническим причинам большинство исследований с фиксацией потенциала осуществлялось на периферических тканях синусового узла вблизи пограничного гребня, где наиболее отрицательный максимальный диастолический потенциал регистрировался в диапазоне от —70 до —75 мВ. Спонтанная деполяризация клеток центральной части узла, по-видимому, зависит в основном от активности медленных каналов, в периферических же клетках эта зависимость является менее определяющей. Частота спонтанной деполяризации пейсмекерных клеток зависит от величины максимального диастолического потенциала, скорости или наклона фазы 4 деполяризации потенциала действия, порогового уровня потенциала, скорости нарастания и амплитуды потенциала в фазу 0, а также от длительности потенциала действия (рис. 6.1). Поэтому снижение частоты спонтанных возбуждений синусового узла может быть обусловлено увеличением максимального диастолического потенциала, уменьшением наклона диастолической деполяризации, менее отрицательным, чем в норме, пороговым потенциалом, уменьшением наклона и амплитуды в фазу 0 или увеличением длительности потенциала действия. Как и при деполяризации в фазу 4, другие характеристики потенциала действия клеток синусового узла определяются зависящей от времени или потенциала флюктуацией мембранной проницаемости для различных ионов. Фаза 0 потенциала действия, по-видимому, зависит от активации как быстрого натриевого тока, так и следующего за ним медленного тока, причем доминирующий ток определяется уровнем потенциала в начале фазы регенерации [29—32]. Поддержание мембранного потенциала покоя, вероятно, зависит от электрогенного натриевого насоса и изменений калиевого потока [17, 33—35], хотя вопрос о точных^ механизмах все еще не решен. Длительность потенциала действия, по-видимому, зависит от характеристик тока, проходящего по медленному каналу, а также от калиевой проводимости. Явное замедление автоматизма синусового узла, электрокардиографически не отличимое от нарушений пейсмекерной функции синусового узла, может быть результатом замедления проведения в области синоатриального соединения. Угнетение синоатриального проведения верапамилом в препаратах синусового узла кролика свидетельствует о том, что медленные каналы существенно влияют на характеристики проведения в околоузловой зоне [36].
Рис. 6.1. Типичный потенциал действия синусового узла. По вертикальной оси — шкала потенциалов (в милливольтах). В точках а и б определяется различная частота возбуждения синусового узла в зависимости от наклона деполяризации в фазе 4. При потенциалах действия б и г с одинаковым наклоном деполяризации в фазе 4 максимальный диастолический потенциал в точке б выше (более отрицательный, чем в точке г); следовательно, частота возбуждения синусового водителя ритма в точке г ниже, чем в точке б. Другой характеристикой, влияющей на частоту возбуждения синусового узла, является величина порогового потенциала (ПП). В точках а, б и г пороговый потенциал составляет приблизительно —70 мВ (ПП-1), а в точке в—примерно —48 мВ (ПП-2), Такой сдвиг порогового потенциала вверх (т. е. к менее отрицательной величине) обусловливает снижение частоты возбуждения (при ее сравнении в точках а и в).
|
||||
|
|
Последнее изменение этой страницы: 2016-09-20; просмотров: 262; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.12.123.41 (0.01 с.) |