Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Элементарные двигательные расстройстваСодержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
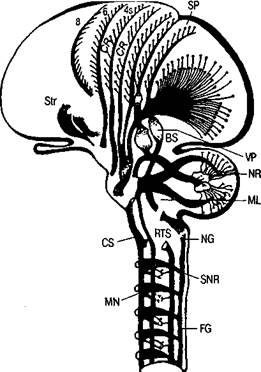
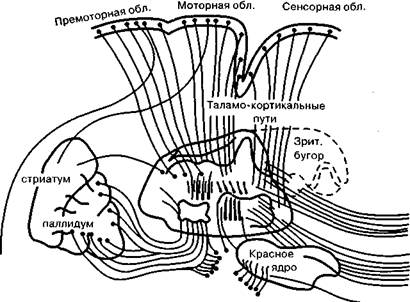
Произвольные движения и действия (как совокупность произвольных движений, объединенных единой целью) относятся к числу наиболее сложных психических функций человека. Их морфофизиологической основой являются сложные функциональные системы — иерархически организованные, включающие много уровней и подуровней, характеризующиеся сложным и многозвенным афферентным и эфферентным составом, условно-рефлекторные по своему происхождению, формирующиеся полностью прижизненно, как и другие высшие психические функции. Современные психологические представления о произвольных движениях сложились в борьбе как с идеалистическим подходом, так и с вульгарно-материалистическими, механистическими концепциями (бихевиоризмом и др.). Важный вклад в современное понимание произвольных движений внесли отечественные физиологи (И. М. Сеченов, И. П. Павлов, Н. А. Бернштейн, П. К. Анохин и др.) и психологи (Л. С. Выготский, А. Н. Леонтьев, А. Р. Лурия, А. В. Запорожец и мн. др.). Произвольные движения и действия могут быть как самостоятельными двигательными актами, так и средствами, с помощью которых реализуются самые различные формы поведения. Произвольные движения входят в состав устной и письменной речи, а также многих других высших психических функций. Произвольные движения включены в разнообразные моторные акты человека, составляя лишь определенную часть (уровень) внутри них. С физиологической точки зрения к произвольным движениям относятся движения поперечно-полосатой мускулатуры рук, лица ног, всего туловища, т. е. обширнейший класс движений. Современная физиология располагает разнообразными сведениями относительно большой сложности как афферентного, так и эфферентного механизмов произвольных движений. О большой сложности и многообразии афферентных механизмов произвольных движений писали И. М. Сеченов, И. П. Павлов и многие другие отечественные физиологи, показавшие, что произвольные движения — это сложно афферентированные системы, включающие разные виды афферентации, среди которых базальной является кинестетическая афферентация. Тем самым была опровергнута вульгарно-материалистическая точка зрения на природу произвольных движений как чисто эффекторных процессов, возникающих Хомская Е. Д. Х = Нейропсихология: 4-е издание. — СПб.: Питер, 2005. — 496 с: ил. 99 вследствие активации только моторных клеток коры больших полушарий (клеток Беца) и мотонейронов спинного мозга. На принципиально важную роль афферентации в регуляции произвольных движений и действий указывали Н. А. Бернштейн и П. К. Анохин, внесшие огромный вклад в развитие материалистического понимания генеза произвольного акта и тем самым — в общую теорию произвольных актов. Н. А. Бернштейн показал принципиальную невозможность реализации произвольного двигательного акта с помощью только одних эфферентных импульсов. Концепция Н. А. Бернштейна (1947) о построении движений имела огромное значение для создания теории произвольного двигательного акта. Согласно данной концепции, любое движение — сложная многоуровневая система, где каждый уровень (или определенные анатомические структуры) характеризуется «ведущей афферентацией» и собственным набором регулируемых движений. Н. А. Бернштейном выделены пять уровней регуляции движений: ♦ рубро-спинальный; ♦ таламо-паллидарный; ♦ пирамидно-стриальный; ♦ теменно-премоторный; ♦ корковый «символический». Все эти уровни объединяют непроизвольные и произвольные движения в единую систему. Первый и второй уровни ответственны за регуляцию непроизвольных движений (к ним относятся движения гладкой мускулатуры, тремор, тонус, синергии, автоматизмы и др.). Третий-пятый уровни связаны с регуляцией произвольных двигательных актов, в которых участвуют как движения всего туловища (ходьба, бег, прыжки и др.), так и отдельных частей тела: рук (действия с предметами, письмо, рисование, различные мануальные навыки), лица (мимика), речевого аппарата (устная речь) и т. д. Таким образом, согласно Н. А. Бернштейну, произвольные движения — это целый набор различных двигательных актов, регулируемых разными уровнями (структурами) нервной системы и управляемых разного рода афферентными импульсами (и различной «ведущей афферентацией»). Поражение любого из перечисленных уровней ведет к нарушениям движений данного уровня, а также тех двигательных актов, куда эти движения включаются как «фоновые». Тип афферентации, а также соответствующие анатомические структуры являются критериями для выделения класса движений (это относится как к произвольным, так и к непроизвольным движениям). Иными словами, афферентация является важнейшим фактором, определяющим тип движения. На принципиальную важность афферентации в регуляции всех поведенческих актов животных (куда и входят так называемые произвольные движения — по терминологии И. П. Павлова) указывал П. К. Анохин, разработавший концепцию функциональных систем. Конечный двигательный акт предопределяется и афферентным синтезом («предпусковой афферентацией»), и текущей афферентацией, поступающей от двигающегося органа, и подкреплением («обратной афферентацией»), без которых полезный результат не может быть достигнут (П. К. Анохин, 1968, 1971 и др.). А. Р. Лурия, анализируя наблюдения над больными с локальными поражениями головного мозга, описал конкретный состав корковых зон, участвующих в мозговой организации произвольных двигательных актов, включив в понятие «двигательный организатор» не только моторные, но и сенсорные, и ассоциативные корковые поля. В статье «Двигательный анализатор и проблема корковой организации движений» (1957) А. Р. Лурия отмечал, что помимо собственно двигательных, моторных зон коры больших полушарий в корковое звено двигательного анализатора следует включать и многие другие зоны коры, а именно: ♦ постцентральную теменную кору, обеспечивающую анализ кожно-кинестетической афферентации, поступающей от органов движения; Из произведений А. Р. Лурия Психологическая наука давно отбросила положение, согласно которому произвольные движения являются недетерминированными волевыми актами, не имеющими афферентной основы. Существенную роль в этом сыграли влияние материалистической философии и те конкретные данные, которые были получены физиологическими исследованиями. И. П. Павлов, опираясь на данные Н. И. Красногорского (1911), Ю. М. Конорского и С. М. Миллера (1936), впервые включил в физиологические основы произвольного движения процессы анализа и синтеза кинестетических сигналов и выдвинул понятие «двигательного анализатора». Тем самым он распространил те принципы, в свете которых рассматривались все афферентные зоны коры головного мозга (кожно-кинестетическая, зрительная, слуховая), на двигательную кору и сделал произвольные движения доступными для объективного физиологического изучения. Работы Н. А. Бернштейна (1926, 1935, 1947 и др.), посвященные исследованию двигательных актов, уточнили их афферентную основу и установили роль обратной афферентации в их управлении. Однако до настоящего времени остается актуальным изучение афферентной основы нарушения высших форм движения и действия. Таким образом, современные научные представления о рефлекторном строении сенсорных и двигательных процессов подчеркивают их взаимозависимость и дают все основания для того, чтобы рассматривать агностические и апраксические расстройства в теснейшей связи друг с другом. (А. Р. Лурия. Высшие корковые функции человека. — М.: Академический проект, 2000. — С. 80.) ♦ задние затылочные и теменно-затылочные отделы коры больших полушарий, которые обеспечивают регуляцию движений с помощью зрительной афферентации, а также ответственны за пространственную Хомская Е. Д. Х = Нейропсихология: 4-е издание. — СПб.: Питер, 2005. — 496 с: ил. 100 организацию движений; ♦ височную кору (прежде всего левого полушария), обеспечивающую не только слухоречевую афферентацию речевой моторики, но и участвующую во всех «оречевленных» (внешней и внутренней речью) двигательных актах; ♦ передние отделы коры больших полушарий (премоторную и префронтальную кору), с помощью которых осуществляются программирование движений, организация движений во времени и контроль за выполнением программы. Следовательно, согласно концепции А. Р. Лурия, произвольные движения человека обеспечиваются самыми различными видами афферентации, и поэтому в корковое звено двигательного анализатора, следует включать почти всю кору больших полушарий. В целом, в современной нейрофизиологии и нейропсихологии сложились представления о том, что произвольные движения — это очень сложно афферентированные системы, которые реализуются при участии почти всей коры больших полушарий. Каковы эфферентные механизмы произвольных движений? К эфферентным (исполнительным) механизмам произвольных движений и действий относятся, как известно, две взаимосвязанные, но в определенной степени автономные эфферентные системы: пирамидная и экстрапирамидная, корковые отделы которых составляют единую сенсомоторную зону коры. Пирамидная система. Пирамидная система. Согласно традиционной точке зрения это основной механизм, реализующий произвольные движения; начинается от моторных клеток Беца, находящихся в V слое моторной коры (4-е поле), продолжается в виде корково-спинномозгового, или пирамидного, тракта, который переходит на противоположную сторону в области пирамид и заканчивается на мотонейронах спинного мозга (на 2-м нейроне пирамидного пути), иннервирующих соответствующую группу мышц. К этим представлениям о пирамидной системе как об основном эфферентном механизме произвольных движений добавились новые данные современной анатомии и физиологии. Во-первых, не только 4-е поле является моторным. Это первичное моторное поле коры, разные участки которого связаны с иннервацией различных групп мышц (см. схему «двигательного человечка» У. Пенфилда и Г. Джаспера на рис. 11). Как известно, первичное моторное поле коры больших полушарий характеризуется мощным развитием V слоя, содержащего самые крупные клетки головного мозга человека (клетки Беца). Эти клетки («гигантские пирамиды») обладают специфическим строением и имеют самый длинный аксон в нервной системе человека (его длина может достигать двух метров), заканчивающийся на мотонейронах спинного мозга. Моторные клетки пирамидного типа обнаружены не только в 4-м поле, но и в 6-м и 8-м полях прецентральной коры, и во 2, 1 и даже в 3-м полях постцентральной коры (и в ряде других областей коры). Следовательно, пирамидный путь начинается не только от 4-го поля, как это пред- полагалось ранее, а со значительно больших площадей коры больших полушарий. По данным П. Дууса (1997), только 40 % всех волокон пирамидного пути начинается в 4-м поле, около 20 % — в постцентральной извилине; остальные — в премоторной зоне коры больших полушарий. Раздражение 4-го поля вызывает сокращение соответствующих групп мышц на противоположной стороне тела. Иными словами, 4-е поле построено по соматотопическому принципу. Во-вторых, установлено, что пирамидный путь содержит волокна различного типа (по диаметру и степени миелинизации). Хорошо миелинизированные волокна составляют не более 10 % всех пирамидных волокон, которые идут от коры к периферии. По-видимому, с их помощью осуществляется фазический (непосредственно исполнительный) компонент произвольных движений. Подавляющее большинство слабо миелинизированных волокон пирамидного пути имеет, вероятно, иные функции и регулирует прежде всего тонические (фоновые, настроечные) компоненты произвольных движений. В-третьих, если ранее предполагалось, что существует единый пирамидный, или кортико-спинальный, путь (латеральный), который идет с перекрестом в зоне пирамид от коры больших полушарий к мотонейронам спинного мозга, то в настоящее время выделен другой кортико-спинальный путь (вентральный), идущий без перекреста в составе пирамид на той же стороне. Эти два пути имеют различное функциональное значение. Наконец, пирамидный путь оканчивается не непосредственно на мотонейронах, расположенных в передних рогах спинного мозга, как считалось ранее, а главным образом на промежуточных (или вставочных) нейронах, с помощью которых модулируется возбудимость основных мотонейронов и тем самым оказывается воздействие на конечный результат — произвольные движения. Все эти данные свидетельствуют о сложности пирамидной системы как исполнительного механизма (рис. 31). Следует отметить, что кроме 4-го моторного поля (о моторных функциях которого известно уже очень давно, после опытов Г. Фритча и Е. Гитцига, раздражавших у животных электрическим током эту зону мозга) у человека обнаружен еще целый ряд моторных зон, при стимуляции которых также возникают двигательные эффекты. Эти зоны, получившие название дополнительных моторных зон, описаны Г. Джаспером, У. Пенфилдом и другими крупнейшими физиологами. Хомская Е. Д. Х = Нейропсихология: 4-е издание. — СПб.: Питер, 2005. — 496 с: ил. 101 Существуют две основные дополнительные моторные зоны коры. Одна из них находится вдоль края Сильвиевой борозды; ее стимуля- Рис. 31. Строение пирамидной и экстрапирамидной систем: FG—NG—ML—VP—SP — проводящие пути и переключения кожно-кинестетических рецепций; CS—BS—NR — связи коры мозга с мозжечком; CR—MN — корково-двигательный пирамидный путь; CR—NR—RTS — связи коры с ядрами ретикулярной формации; CR—SNR — связи коры с черной субстанцией; Str— подкорковые узлы больших полушарий (по Д.
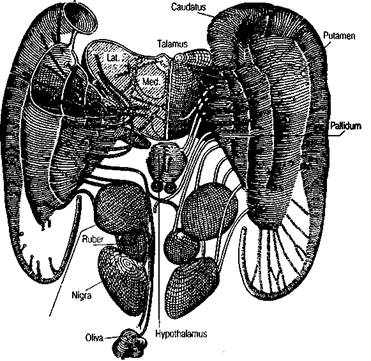
ция вызывает движения рук и ног (как ипсилатеральных, так и контралатеральных). Другая расположена на внутренней медиальной поверхности полушарий кпереди от моторной зоны, в прецентральных отделах мозга. Раздражение этой зоны также приводит к различным двигательным актам. Важно отметить, что при раздражении дополнительных моторных зон возникают не элементарные сокращения отдельных мышечных групп, а целостные комплексные движения, что говорит об их особом функциональном значении (У. Пенфилд, Г. Джаспер, 1958). Описаны и другие дополнительные моторные зоны коры. Все эти данные говорят о том, что современные знания о корковой организации произвольных движений еще далеко не полны. Помимо моторных зон коры больших полушарий, раздражение которых вызывает движения, существуют и такие зоны коры, раздражение которых прекращает уже начавшееся движение (подавляющие области коры). Они расположены кпереди от 4-го поля (поле 4s) на границе 4-го и 6-го полей; кпереди от 8-го поля (поле 8s); кзади от 2-го поля (поле 2s) и кпереди от 19-го поля (поле 19s). На внутренней поверхности полушария находится подавляющее поле 24s (см. рис. 4). Поля 8s, 4s, 2s и 19s — это узенькие полоски, разграничивающие основные области коры, связанные с проекциями ядер зрительного бугра на кору больших полушарий. Как известно, задние отделы коры (17, 18, 19-е поля) являются зоной проекции латерального коленчатого тела; сенсомоторная область — зона проекции вентральных таламических ядер; префронтальная область — зона проекции ДМ-ядра зрительного бугра. Таким образом, подавляющие полоски коры разграничивают сферы влияния разных реле-ядер таламуса. В коре больших полушарий имеются еще особые адверзивные зоны. Это области коры, хорошо известные нейрохирургам и невропатологам. Их раздражение (электрическим током или болезненным процессом) вызывает адверзивные эпилептические припадки (начинающиеся с адверзии — поворота туловища, глаз, головы, рук и ног в сторону, противоположную расположению возбуждающего агента). Эпилепсия, протекающая с припадками этого типа, известна как «эпилепсия Джексона». Существуют две адверзивные зоны коры: премоторная и теменно-затылочная (поля 6, 8 и 19-е на границе с 37-м, 39-м полями). Предполагается, что эти поля коры участвуют в сложных реакциях, связанных с вниманием к раздражителю, т. е. в организации сложных двигательных актов, опосредующих внимание к определенному стимулу. Хомская Е. Д. Х = Нейропсихология: 4-е издание. — СПб.: Питер, 2005. — 496 с: ил. 102 Экстрапирамидная система Экстрапирамидная система — второй эфферентный механизм реализации произвольных движений и действий. Экстрапирамидной системой называются все двигательные пути, которые не проходят через пирамиды продолговатого мозга (рис. 32; цветная вклейка). Как уже говорилось выше, пирамидная система не является единственным механизмом, с помощью которого осуществляются произвольные двигательные акты. Экстрапирамидная система более древняя, чем пирамидная. Она очень сложна по своему составу. До настоящего времени идут споры о том, какие именно структуры относить к экстрапирамидной системе и, главное, каковы функции этих структур. В экстрапирамидной системе различают корковый и подкорковый отделы. К корковому отделу экстрапирамидной системы относятся те же поля, которые входят в корковое ядро двигательного анализатора. Это 6-е и 8-е, а также 1-е и 2-е поля, т. е. сенсомоторная область коры. Исключение составляет 4-е поле, которое является корковым звеном только пирамидной системы. 6-е и 8-е поля по строению принадлежат к полям моторного типа, в III и V слоях этих полей расположены пирамидные моторные клетки, но меньшего размера, чем клетки Беца. Строение подкоркового отдела экстрапирамидной системы довольно сложное. Он состоит из целого ряда образований. Прежде всего это стриопаллидарная система — центральная группа образований внутри экстрапирамидной системы, куда входят хвостатое ядро, скорлупа и бледный шар (или паллидум). Эта система базальных ядер располагается внутри белого вещества (в глубине премоторной зоны мозга) и характеризуется сложными двигательными функциями (рис. 33). Следует отметить, что, по современным данным, стриопаллидарная система участвует в осуществлении не только моторных, но и ряда других сложных функций (см. гл. 22). В экстрапирамидную систему входят и другие образования: поясная кора, черная субстанция, Льюисово тело, передневентралъные, интраламинарные ядра таламуса, субталамические ядра, красное ядро, мозжечок, различные отделы ретикулярной формации, ретикулярные структуры спинного мозга. Конечной инстанцией экстрапирамидных влияний являются те же мотонейроны спинного мозга, к которым адресуются импульсы и пирамидной системы (рис. 34). Четкая анатомическая граница между пирамидной и экстрапирамидной системами отсутствует (рис. 35). Они обособлены анатомически только на участке пирамид, в продолговатом мозге. Однако функциональные различия между этими системами достаточно отчетливы. И особенно ясно они проявляются в клинике локальных поражений головного мозга. Нарушения двигательных функций, возникающие при различных локальных поражениях мозга, можно подразделить на относительно элементарные, связанные с поражением исполнительных, эфферентных механизмов движений, и более сложные, распространяющиеся на произвольные движения и действия и связанные преимущественно с поражением афферентных механизмов двигательных актов. Хомская Е. Д. Х = Нейропсихология: 4-е издание. — СПб.: Питер, 2005. — 496 с: ил. 103 Рис. 33. Стриопаллидум и его эфферентные связи, базальный аспект. Хвостатое ядро (с. caudatus), скорлупа (putamen) и бледный шар (pallidum), составляющие единую систему (striapallidum), тесно связаны с глубокими структурами: зрительным бугром, гипоталамической областью, красным ядром, черной субстанцией и корой головного мозга (по О. Криигу)
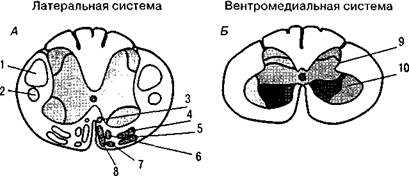
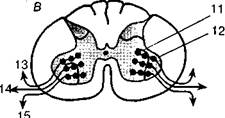
Subthalamic Относительно элементарные двигательные расстройства возникают при поражении подкорковых звеньев пирамидной и экстрапирамидной систем. При поражении коркового звена пирамидной системы (4-го поля), расположенного в прецентральной области, наблюдаются двигательные расстройства в виде парезов или параличей определенной группы мышц: руки, ноги или туловища на стороне, противоположной поражению. Для поражения 4-го поля характерен вялый паралич (когда мышцы не сопротивляются пассивному движению), протекающий на фоне снижения мышечного тонуса. Но при очагах, расположенных кпереди от 4-го поля (в 6-м и 8-м полях коры), возникает картина спастического паралича, т. е. выпадений соответствующих движений на фоне повышения мышечного тонуса. Явления паре- Хомская Е. Д. Х = Нейропсихология: 4-е издание. — СПб.: Питер, 2005. — 496 с: ил. 104 Рис. 34. Организация моторных функций на спинальном уровне: А — латеральный и вентромедиальный пути; Б — окончание латерального и вентрального путей на мотонейронах передних отделов спинного мозга и на промежуточных нейронах промежуточной зоны; В — проекция промежуточных нейронов на мотонейроны и проекция мотонейронов на мускулатуру тела. Латеральная система проецируется на дистальную мускулатуру, а вентро-медиальная — на проксимальную. 1 — латеральный кортико-спинальный тракт; 2 — рубро-спинальный тракт; 3 — медиальный тракт; 4 — латеральный ретикуло- спинальный тракт; 5 — вестибуло-спинальный тракт; 6 — медиальный ретикуло-спинальный тракт; 7 — текто-спинальный тракт; 8 — вентральный кортико-спинальный тракт; 9 — промежуточная зона; 10 — вентральная зона; 11 —промежуточные (вставочные) нейроны; 12 — мотонейроны; 13 — пальцы; 14 — руки; 15 — тело (по Л. Лауренсу и Д. Куперсу)
зов вместе с чувствительными расстройствами характерны и для поражения постцентральных отделов коры. Эти нарушения двигательных функций подробно изучаются неврологией. Наряду с этими неврологическими симптомами поражение коркового звена экстрапирамидной системы дает также нарушения сложных произвольных движений, о которых речь будет идти ниже. Рис. 35. Различные системы афферентаций чувствительных (кинестетических) и двигательных (кинетических) отделов коры (по Д. Пейпецу)
При поражении пирамидных путей в подкорковых областях мозга (например, в зоне внутренней капсулы) возникает полное выпадение движений (паралич) на противоположной стороне. Полное одностороннее Хомская Е. Д. Х = Нейропсихология: 4-е издание. — СПб.: Питер, 2005. — 496 с: ил. 105 выпадение движений руки и ноги (гемиплегия) появляется при грубых очагах. Чаще в клинике локальных поражений мозга наблюдаются явления частичного снижения двигательных функций на одной стороне (гемипарезы). При пересечении пирамидного пути в зоне пирамид — единственной зоне, где пирамидный и экстрапирамидный пути анатомически обособлены, — произвольные движения реализуются только с помощью экстрапирамидной системы. В 40-е годы XX века эксперименты по пересечению пирамид проводились английским физиологом С. Тоуэром на обезьянах. Оказалось, что обезьяна способна выполнять целый ряд сложных движений (захватывания, манипуляции с предметами и др.). Однако в подобных случаях движения становятся менее дискретными (т. е. более грубыми). В захватывании предметов начинает участвовать большее количество мышц. Кроме того, в движени- ях, где требуется участие всех мышц (типа прыжка), что характерно для моторики обезьян, исчезают точность и координированность. Подобные нарушения движений протекают на фоне повышения мышечного тонуса. Эти данные свидетельствуют о том, что пирамидная система участвует в организации преимущественно точных, дискретных, пространственно-ориентированных движений и в подавлении мышечного тонуса. Поражение корковых и подкорковых звеньев экстрапирамидной системы приводит к появлению различных двигательных расстройств. Эти расстройства можно подразделить на динамические (т. е. нарушения собственно движений) и статические (т. е. нарушения позы). При поражении коркового уровня экстрапирамидной системы (6-е и 8-е поля премоторной коры), который связан с вентролатеральным ядром таламуса, бледным шаром и мозжечком, в контралатеральных конечностях возникают спастические двигательные нарушения. Раздражение 6-го или 8-го полей вызывает повороты головы, глаз и туловища в противоположную сторону (адверзии), а также сложные движения контралатеральных руки или ноги. Поражение подкорковой стриопаллидарной системы, вызванное различными заболеваниями (паркинсонизмом, болезнью Альцгеймера, Пика, опухолями, кровоизлияниями в область базальных ядер и др.), характеризуется общей неподвижностью, адинамией, трудностями передвижения. Одновременно появляются насильственные движения контралатеральных руки, ноги, головы — гиперкинезы. У таких больных наблюдаются и нарушение тонуса (в виде спастичности, ригидности или гипотонии), составляющего основу позы, и нарушение двигательных актов (в виде усиления тремора — гиперкинезов). Больные теряют возможность себя обслуживать и становятся инвалидами. В настоящее время разработаны хирургические (стереотаксические) и консервативные (фармакологические) методы лечения паркинсонизма. Суть первых состоит в разрушении одной из «больных» экстрапирамидных структур (например, вентролатерального ядра таламуса) и прекращении патологического возбуждения, циркулирующего по замкнутым экстрапирамидным путям. В зависимости от формы паркинсонизма разрушают также либо зону паллидума, либо зону стриатума (в области хвостатого ядра или скорлупы). Зона паллидума (бледный шар) имеет отношение к регуляции медленных плавных движений и к поддержанию тонуса (позы), в то время как зона стриатума (скорлупа и хвостатое ядро) в большей степени связана с физиологическим тремором, переходящим при патологии в гиперкинезы. Избирательное поражение зоны паллидума (более древней части, чем стриатум) может привести к атетозу или хореоатетозу (патологическим волнообразным движениям рук и ног, подергиванию конечностей и т. д.). Эти заболевания хорошо изучены и описаны в неврологических руководствах. Поражение стриопаллидарных образований сопровождается еще одним видом двигательных симптомов — нарушением мимики и пантомимики, т. е. непроизвольных моторных компонентов эмоций. Эти нарушения могут выступать либо в форме амимии (маскообразное лицо) и общей обездвиженности (отсутствие непроизвольных движений всего тела при различных эмоциях), либо в форме насильственного смеха, плача или насильственной ходьбы, бега (пропульсии). Нередко у этих больных страдает и субъективное 'переживание эмоций. Наконец, у таких больных нарушаются и физиологические синергии — нормальные сочетанные движения разных двигательных органов (например, размахивание руками при ходьбе), что приводит к неестественности их двигательных актов. Последствия поражения других структур экстрапирамидной системы изучены в меньшей степени, за исключением, конечно, мозжечка. Мозжечок представляет собой важнейший центр координации различных двигательных актов, «орган равновесия», обеспечивающий целый ряд безусловных моторных актов, связанных со зрительной, слуховой, кожно-кинестетической, вестибулярной афферентацией. Поражение мозжечка сопровождается разнообразными двигательными расстройствами (прежде всего расстройствами координации двигательных актов). Их описание составляет один из хорошо разработанных разделов современной неврологии. Поражение пирамидных и экстрапирамидных структур спинного мозга сводится к нарушению функций мотонейронов, вследствие чего выпадают (или нарушаются) управляемые ими движения. В зависимости от уровня поражения спинного мозга нарушаются двигательные функции верхних или нижних конечностей (на одной или на обеих сторонах), причем все местные двигательные рефлексы осуществляются, как правило, Хомская Е. Д. Х = Нейропсихология: 4-е издание. — СПб.: Питер, 2005. — 496 с: ил. 106 нормально или даже усиливаются вследствие устранения коркового контроля. Все эти нарушения движений также подробно рассматриваются в курсе неврологии. Клинические наблюдения за больными, у которых имеется поражение того или иного уровня пирамидной или экстрапирамидной системы, позволили уточнить функции этих систем. Пирамидная система ответственна за регуляцию дискретных, точных движений, полностью подчиненных произвольному контролю и хорошо афферентированных «внешней» афферентацией (зрительной, слуховой). Она управляет комплексными пространственно-организованными движениями, в которых участвует все тело. Пирамидная система регулирует преимущественно фазический тип движений, т. е. движения, точно дозированные во времени и в пространстве. Экстрапирамидная система управляет в основном непроизвольными компонентами произвольных движений; к ним кроме регуляции тонуса (того фона двигательной активности, на котором разыгрываются фазические кратковременные двигательные акты) относятся: ♦ поддержание позы; ♦ регуляция физиологического тремора; ♦ физиологические синергии; ♦ координация движений; ♦ общая согласованность двигательных актов; ♦ их интеграция; ♦ пластичность тела; ♦ пантомимика; ♦ мимика и т. д. Экстрапирамидная система управляет также разнообразными двигательными навыками, автоматизмами. В целом экстрапирамидная система менее кортиколизована, чем пирамидная, и регулируемые ею двигательные акты в меньшей степени произвольны, чем движения, регулируемые пирамидной системой. Следует, однако, помнить, что пирамидная и экстрапирамидная системы представляют собой единый эфферентный механизм, разные уровни которого отражают разные этапы эволюции. Пирамидная система, как эволюционно более молодая, является в известной степени «надстройкой» над более древними экстрапирамидными структурами, и ее возникновение у человека обусловлено прежде всего развитием произвольных движений и действий.
|
||||
|
|
Последнее изменение этой страницы: 2016-08-25; просмотров: 490; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.147.89.50 (0.012 с.) |