Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Экологические группы гетеротрофных высших растенийСодержание книги
Похожие статьи вашей тематики
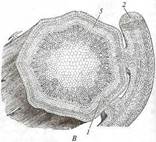
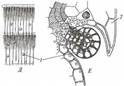
Поиск на нашем сайте Паразиты полностью или частично существуют за счет автотрофных растений. На растениях паразитируют только спорофиты некоторых покрытосеменных и одного вида голосеменных растений. Среди них различают полупаразитов, самостоятельно обеспечивающих себя ассимилятами, но извлекающих из растений-хозяев воду с элементами минерального питания (рис. 233), и голопаразитов, существующих за счет хозяина и лишенных хлорофилла. Корневые паразиты, к которым принадлежит большинство паразитических семенных растений, извлекают ресурсы из корней растений-хозяев с помощью гаусторий. Корневые полупаразиты (рис. 233, А — В) имеют нормально развитую побеговую систему, но слабо развитую корневую, в которой остались только скелетные корни, а вместо сосущих корней образуются гаустории. Корневые голопаразиты (рис. 234, Г—Е) большую часть жизни проводят на корнях хозяина в почве в виде протокорм - клубневидного, не дифференцированного на органы тела, присосавшегося гаусторией к корню хозяина. В сезон размножения на протокорме образуются надземные побеги с соцветием и редуцированными листьями.
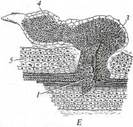
Стеблевые паразиты используют для питания стебли растений-хозяев. Стеблевые полупаразиты поселяются в кроне дерева-хозяина подобно эпифитам (см. рис. 233, Г, Д). Однако в отличие от последних их скелетные корни растут внутри ветви хозяина, между ксилемой и флоэмой. Сосущие корни, как и у корневых полупаразитов, замещены гаусториями. У стеблевых голопаразитов (см. рис. 234, А— В) гипокотиль, главный корень и основание побега отмирают вскоре после прорастания семени, а побег с длинными междоузлиями и очень мелкими чешуевидными листьями обвивает стебель хозяина. Гаустории образуются в любом месте стебля голопаразита, где он тесно соприкасается со стеблем хозяина. Все вышеописанные паразиты - эктопаразиты, большая часть их вегетативного тела находится рядом с телом хозяина, в которое они проникают только гаусториями или - редко - корнями. Среди двудольных растений есть несколько эндопаразитов, все вегетативное тело которых развивается внутри тела хозяина (рис. 235). Оно представлено ветвящимися одно-двурядными цепочками клеток, очень напоминающими гифы грибов. Местами у поверхности хозяина на «гифах» эндопаразита развиваются протокормы, на которых эндогенно образуются зачатки соцветий или одиночных цветков. Развивающиеся соцветия и бутоны прорывают наружные ткани протокорма и хозяина,
вынося цветки наружу. Все эндопаразитические виды голопаразиты: одни заселяют стебли хозяина, а другие — преимущественно корни. Микотрофные паразиты. Очень многие виды высших растений живут в симбиозе с грибами по типу мутуализма, когда оба организма извлекают выгоду от сожительства. Мутуалистические отношения гриба и растения представляют собой сбалансированный умеренный взаимный паразитизм. Некоторые растения в процессе эволюции перешли к одностороннему временному или постоянному паразитизму на грибах. Для целых семейств цветковых растений (орхидные, вересковые) характерен паразитизм проростков на грибах. Их семена прорастают только после внедрения в их ткани гиф гриба, а проростки довольно долго (до нескольких лет) ведут подземный образ жизни, частично переваривая гифы гриба.
Впоследствии они развивают нормальные олиственные зеленые надземные побеги и от паразитизма на грибе переходят к мутуалистическим взаимоотношениям с ним. Однако растения некоторых видов на всю жизнь остаются паразитами гриба (рис. 236). Они либо вообще лишены корней, либо имеют короткие коралловидные корни, заселенные грибом. Их побеговые системы сходны с таковыми корневых голопара-зитов. К микотрофным голопаразитам относятся и гаметофиты многих плаунов и некоторых папоротников, представленные подземными бесхлорофилльными протокормами. Паразитизм растений на грибах настолько резко контрастирует с привычным паразитизмом грибов на растениях, что ботаники долго отказывались признавать микотрофные паразиты и считали их сапрофитами, потребляющими разлагающееся органическое вещество почвы. Однако у растений нет ферментных систем для разложения отмершей органики, и потому они не могут быть сапротрофами. Ключевое значение для выживания любых паразитов имеет их встреча с хозяином. Поскольку высшие растения не способны активно разыскивать и преследовать хозяина, то паразиты образуют огромное число семян. Это гарантирует то, что некоторые из них окажутся рядом с хозяином, и проросший паразит будет обеспечен ресурсами. Хищные растения полностью обеспечивают себя фотоассимилятами, но ловят животных для дополнительного азотного питания. Хищные растения никогда не используют корни для ловли и переваривания добычи, их ловчие структуры всегда развиваются на побегах (см. рис. 237 - 239). В простейшем случае это ловчие трихомы или эмергенцы на листьях (иногда на стеблях), выделяющие клейкую слизь (рис. 237). Прилипших к ним мелких животных растение переваривает с помощью выделяемого особыми железками секрета, сходного с пепсином желудочного сока. Длинные ловчие эмергенцы сгибаются, прижимая жертву к пищеварительным железкам. У некоторых видов лист заворачивается вокруг пойманного животного, что интенсифицирует его перс-варивание.
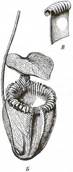
Более сложную ловчую систему представляют асцидиатные листья (рис. 238, см. также рис. 77, А). По краю устья асцидиатной части листа развивается нектарник, часто ярко окрашенный. Вероятно, выделяемый нектар содержит также какие-то вещества, нарушающие координацию движений у насекомых. Насекомые по неосторожности сваливаются внутрь асцидия и не могут выбраться из-за того, что устье его в виде карниза нависает над полостью асцидия, а стенки асцидия внутри густо покрыты уплощенными, обращенными базипетально трихомами, допускающими движение только ко дну. В нижней части асцидий заполнен жидкостью, в которой тонут пойманные животные. Пищеварительного фермента у многих растений с таким ловчим аппаратом, по-видимому, нет и разложение жертвы осуществляют симбиотические бактерии. На стенках базаль-ной части асцидия есть специальные всасывающие трихомы, поглощающие продукты бактериального разложения жертвы. Над устьем асцидия у многих видов нависает неподвижный клапан, предотвращающий попадание в асцидий дождевой воды. У некоторых видов клапан играет важную роль в поимке добычи. Он содержит участок прозрачной ткани, на который ориентируется насекомое, взлетающее с нектарника. Ударившись о клапан, оно падает в асцидий. Ловчие пузырьки, активно захватывающие добычу, присущи водным хищным растениям из рода пузырчатка (Utricularia). Их строение и принцип работы описаны в разделе 1.5.2. Пищеварительных ферментов у пузырчаток, вероятно, нет и пойманные жертвы погибают от удушья и разлагаются бактериями и простейшими, обитающими в пузырьке. На внутренней поверхности пузырька находятся всасывающие трихомы двух типов. Одни поглощают продукты разложения жертвы, другие откачивают из пузырька воду, подготавливая его, таким образом, к захвату новой жертвы (рис. 239).
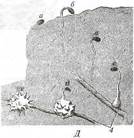
Ловчие части листа венериной мухоловки (Dionaea) и альдрованды (Aldrovanda) представлены двумя подвижными створками, действующими по принципу капкана. На адаксиальной стороне каждой створки находятся жесткие чувствительные трихомы. Смещение такого трихома, задетого животным, генерирует в его базальной клетке разность потенциалов на плаз-малемме. По специальным клеткам этот импульс передается к моторным клеткам на адаксиальной стороне листа. Скорость передачи импульса на порядок меньше скорости передачи нервного импульса по нейрону. Моторные клетки, получив сигнал, резко теряют тургор, вследствие чего створки захлопываются на адаксиальной стороне. Железистые трихомы на адаксиальной стороне створок выделяют протеолитический фермент. После переваривания добычи створки раздвигаются, и лист может ловить очередную жертву. ЖИЗНЕННЫЕ СТРАТЕГИИ ВИДОВ Под жизненными стратегиями понимают обобщенные биологические свойства видов, отражающие специфику их взаимодействия с совокупным действием факторов внешней среды. Рассмотренные в предыдущем разделе экологические группы растений отражают стратегию приспособления к факторам среды, даже самым неблагоприятным. В случае резко выраженной сезонности климата у растений появляется возможность избегать действия самых неблагоприятных факторов среды обитания. Такую стратегию реализуют многие виды высших растений, вегетирующих только в сезон с наиболее благоприятными значениями факторов среды, а в остальное время впадающие в анабиоз. Наиболее ярко стратегия избегания неблагоприятных внешних воздействий выражена у эфемеров (рис. 240), которые за 3-5 недель оптимальных условий среды успевают пройти весь жизненный цикл и отмирают, оставив покоящиеся семена или споры, хорошо защищенные покровами от влияния внешних условий. Такую же стратегию реализуют и эфемероиды (рис. 241), после краткого периода вегетации переживающие неблагоприятный период в форме лишенных надземных побегов, а часто и корней покоящихся луковиц, корневищ, клубнелуковиц и клубней, укрытых в почве. Другой аспект взаимодействия растений с окружающей их средой отражает система стратегий Макартура. Эти стратегии возникают в результате действия соответственно r- и К-отбора. К-отбор направлен на выработку морфолого-функциональных адаптации, обеспечивающих выживание взрослого организма. Развившиеся под его действием организмы имеют высокую индивидуальную устойчивость к внешним воздействиям, низкую смертность. но медленно размножаются. Поэтому растения, обладающие такой стратегией, тратят основную долю ресурсов на рост и функционирование своих органов и значительно меньше расходуют их на размножение. Напротив, г-отбор направлен на интенсификацию размножения, чем достигается устойчивое существование вида при низкой устойчивости отдельных организмов. У видов, сформировавшихся под действием r-отбора, недолговечность отдельных растений сочетается с образованием очень большого потомства. Соответственно растения, выработавшие такую стратегию, основную часть своих вещественно-энергетических ресурсов используют на размножение и меньшую - на развитие собственных вегетативных органов.
Между стратегиями приспособления/избегания неблагоприятных условий среды и стратегиями, описанными Макартуром, нет однозначного соответствия. Так, избегающие неблагоприятных факторов эфемеры в полном виде реализуют стратегию г-отбора, а эфемероиды, вкладывая значительные ресурсы в образование корневищ, луковиц и т.п., демонстрируют стратегию К-отбора. Система стратегий Л. Г. Раменского и практически идентичная ей система CSR-стратегий Дж. Трайма отражают преимущественно специфику взаимодействия между растениями разных видов. Виоленты, или Competitors)-виды, включают конкурентно мощные растения, населяющие богатые ресурсами биотопы. Они реализуют стратегию К-отбора и, потребляя основную долю ресурсов, предопределяют условия жизни растений других видов. Это долговечные быстрорастущие растения, способные долго удерживать занятое местообитание, но плохо переносящие нарушения биотопа. Патиенты, или S(tress-tolerant) - виды, приспособлены существовать в экстремальных условиях, потребляя скудные ресурсы среды. Некоторые геоботаники подразделяют их на экотопические патиенты, населяющие олиготрофные биотопы, и фитоценотические патиенты, довольствующиеся ресурсами, не потребленными виолентами. Патиенты - долговечные медленно растущие растения, реализующие стратегию К-отбора и плохо переносящие нарушения биотопа. Эксплеренты, или R(uderals) - виды, - быстро развивающиеся растения, потребляющие значительные ресурсы, но не способные эффективно конкурировать за них с другими видами. Поэтому они развиваются после нарушения биотопа, пока в нем отсутствуют ви-оленты, или сосуществуют с ними, но потребляют ресурсы местообитания в период, когда они не востребованы виолентами. В первом случае эксплеренты представлены недолговечными растениями, реализующими стратегию г-отбора. Во втором случае они находятся под действием К-отбора, долговечны, но большую часть жизни пребывают в анабиозе, как эфемероиды. Большинство видов сочетает в той или иной форме признаки разных стратегий. Поэтому Грайм к трем группам видов добавил группы CS-, CR-, SR- и CSR-видов, в более или менее равной мере сочетающие свойства двух или всех трех первичных стратегий.
|
||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-08-12; просмотров: 942; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.214 (0.012 с.) |