Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Экологические группы видов по отношению к влажностиСодержание книги
Похожие статьи вашей тематики
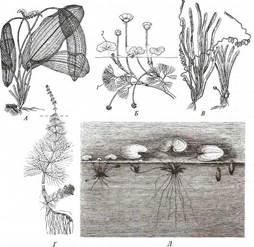
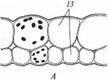
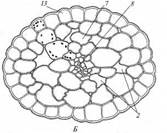
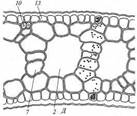
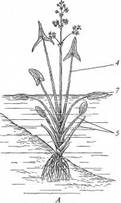
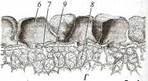
Поиск на нашем сайте Гидрофиты - это растения, обитающие в воде (рис. 222, 223). Данная группа неоднородна, что дает основание различать в се пределах три подгруппы, иногда рассматриваемые как самостоятельные. Отрастающие от дна или свободноплавающие водные растения, все вегетативные органы которых находятся в толще воды, относят к гидатофитам. У некоторых из них репродуктивные органы также развиваются и функционируют под водой, у других репродуктивные органы выставляются в воздушную среду (см. рис. 222, Б). Корневая система у большинства гидатофитов развита слабо, причем корни не имеют корневых волосков. У некоторых гидатофитов корней нет вовсе, и поглощает вещества вся поверхность тела. Так как вегетативные органы гидатофитов не контактируют с воздушной средой, устьиц у них нет. Растения поглощают СО2 в виде растворенного в воде аниона НСО3. Хотя общее содержание углекислоты в воде может значительно превышать фоновую концентрацию СО2 в воздухе, она менее доступна растениям. Это обусловлено тем, что скорость диффузии гидрокарбонатного аниона в 104 раз ниже, чем СО2 - в воздухе, а также образованием на поверхности растения известковой корочки вследствие превращения растворимого гидрокарбоната кальция в нерастворимый карбонат при поглощении из воды НСО3. Вероятно, поэтому ряд гидатофитов поглощает практически всю необходимую для фотосинтеза углекислоту корнями. Кроме того, вода сильно снижает освещенность. Поэтому обитание в толще воды затрудняет использование ресурсов, необходимых для фотосинтеза. В связи с этим для гидатофитов характерна хлорофиллоносная эпидерма и малый удельный объем хлорофиллоносных тканей, который достигается тремя способами: 1) очень тонкими пластинчатыми листьями, 2) нитевидно рассеченными листьями или 3) толстыми листовыми пластинками, основной объем которых занимают воздухоносные полости.
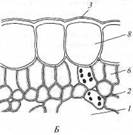
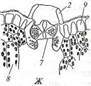
Мощно развитая у гидатофитов аэренхима (см. рис. 223) служит резервуаром, в котором ночью накапливается выделяемый при дыхании тканей СО2, реутилизируемый днем в фотосинтезе. Кроме того, аэренхима быстро транспортирует к ассимилирующим клеткам побега поглощенный корнями СО2 в газообразной форме. По аэренхиме же выделяемый в фотосинтезе О2поступает к корням и корневищам, компенсируя его недостаточное поступление извне вследствие его низкой растворимости в воде и низкого содержания в среде. Аэренхима придает телу растения хорошую плавучесть, благодаря чему оно располагается в пространстве, опираясь на воду. Таким образом, аэренхима у гидатофитов очень своеобразно выполняет функцию специализированной механической ткани. В связи с отсутствием транспирации продольный транспорт воды по телу гидатофитов минимален, и ксилема развита очень слабо. Передвижение воды по ней сопряжено с рекрецией воды через эпидерму или гидатоды. (Гидатодам водных растений часто приписывают способность не только рекретировать воду, но и поглощать ее. На этом оснований их относят к особому типу структур - гидропотам). Аэрогидатофиты имеют листовые пластинки, плавающие по поверхности воды. Их адаксиальная сторона обращена в воздушную среду. В эту группу входят как укореняющиеся на дне растения, так и свободноплавающие у поверхности воды. Репродуктивные органы у большинства видов этой группы располагаются над водой. При большом общем сходстве с гидатофитами аэрогидатофиты имеют ряд специфических черт организации, связанных с наличием контактирующих с воздушной средой плавающих листьев. Многим видам свойственна гетерофиллия вследствие того, что наряду с плавающими листьями у них есть листья, целиком погруженные в воду. Плавающие по поверхности воды пластинки листьев всегда цельные, более или менее округлые, что придает им плавучесть и устойчивость. Их адаксиальная, обращенная вверх сторона не заливается водой благодаря мощной гидрофобной кутикуле. Плавающие листья фотосинтезируют в значительно лучших, чем погруженные листья, условиях освещенности и снабжения СО2, который они поглощают через многочисленные устьица в адаксиальнои эпидерме. Поэтому у аэрогидатофитов развиваются толстые плавающие листовые пластинки с большим удельным объемом хлоренхимы. Клетки эпидермы тоже содержат хлоропласта, но мелкие и немногочисленные и потому в целом не играют существенной роли в фотосинтезе листовой пластинки. У многих видов хлоренхима дифференцирована на палисадную и губчатую. В пластинке развиты крупные межклетники, по которым поглощаемый из воздуха через устьица О2 поступает в аэренхиму для снабжения подводных органов. Кроме того, эти межклетники придают листовой пластинке необходимую плавучесть. Плавающие листья транспирируют, вследствие чего у аэрогидатофитов продольный транспорт воды по растению выше и соответственно ксилема развита значительно сильнее, чем у гидатофитов. Впрочем, транс-пирация в насыщенном водяным паром приводном слое воздуха часто низкая, и на обращенной в воду абаксиальной стороне плавающего листа обычно находятся гидатоды, выделяющие избыточную воду. Реофиты включают виды прикрепленных погруженных растений, обитающие в мелких ручьях и речках с быстрым течением. Условия фотосинтеза для них более благоприятны, чем для гидатофитов, но из-за быстрого течения растения постоянно испытывают резкие динамические нагрузки. Реофиты выработали два способа выживания в таких условиях. У одних располагающиеся в потоке побеги обладают минимальной парусностью вследствие рассечен-ности листьев на тонкие доли. У мадагаскарского апоногетона окончатого (Aponogeton fenestralis) листья с цельными широкоовальными пластинками, но в ходе морфогенеза их ткани в ареолах разрушаются. В результате в листовой пластинке остаются только проводящие пучки, окруженные хлоренхимой, и пластинка принимает вид ажурной сетки с малой парусностью (см. рис. 222, А). Другой способ выживания в быстром потоке реализовали виды из семейства Подостемоновые (Podostemonaceae) (см. рис. 86, Б и 166). Они имеют плотно прилегающие к камням метаморфизированные плоские ассимилирующие корни, напоминающие пластинчатые талломы, а их побеги редуцированы. Некоторые виды вообще не имеют вегетативных побегов и развивают на корнях только придаточные соцветия с очень мелкими цветками и чешуевидными листьями. Гелофиты представляют промежуточную группу между водными и наземными растениями. Они населяют как мелководья, так и регулярно заливаемые и избыточно увлажненные плохо дренированные места. Эти растения укореняются в грунте, а их листья и побеги хотя бы частично возвышаются над водой (рис. 224) (Иногда попавшие на слишком глубокое место гелофиты полностью погружены в воду. Такие растения не способны пройти все стадии жизненного цикла и отмирают, не оставив потомства. Некоторые виды гелофитов также не могут пройти все стадии жизненного цикла и оставить потомство, если развиваются полностью в наземных условиях). Ряду видов свойственна гетерофиллия, причем развиваются листья двух или трех формаций: подводные и надводные или подводные, плавающие и надводные. Подводные и плавающие листья устроены так же, как подобные им листья гидато- и аэрогидатофитов. Надводные листья сходны с листьями наземных растений. Их листовые пластинки толстые, кожистые, гипостомные. Основные клетки эпидермы содержат лейкопласты и не участвуют в фотосинтезе. Мезофилл выполнен хлоренхимой, дифференцированной на адаксиальную палисадную зону плотно расположенных клеток и абаксиальную губчатую зону, в которой находятся очень крупные межклетники (воздухоносные ходы). Межклетники связаны с мощно развитой аэренхимой стебля и корней, по которой поглощаемый через устьица О2 транспортируется к корневищу и корням, не способным непосредственно получить необходимое им количество кислорода из-за более или менее выраженной аноксичности грунта.
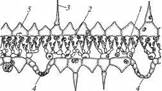
Надводные листья интенсивно транспирируют. Поэтому у гелофитов хорошо развита ксилема, способная пропускать большое количество воды. Тем не менее, гелофиты, особенно тропические и субтропические, часто испытывают значительный суточный дефицит воды, вызванный резким усилением транспирации из-за сильного разогрева воздуха в послеполуденные часы. Соответственно в структуре листьев гелофитов хорошо выражены приспособления для снижения транспирации: толстая кутикула, несколько погруженные устьица. У некоторых видов адаксиальная эпидерма сложена крупными сильно обводненными клетками, что уменьшает интенсивность проникающего в лист света и создает определенный запас воды на время ее дефицита. У ряда видов фотосинтез осуществляют стебли, а листья редуцированы до чешуи. В стеблях и листьях гелофитов хорошо развиты механические элементы, обеспечивающие им необходимую прочность для сохранения определенного положения в пространстве. Некоторые виды обладают пневматофорами (см. рис. 82, 160). У растений мангровы развиваются ходульные корни (см. рис. 83), благодаря которым крона оказывается выше уровня полной воды в прилив. Гигрофиты - наземные растения, населяющие биотопы, в которых влажность воздуха постоянно близка к 100 %, а почва насыщена капиллярной водой, но из-за хорошей дренированности в ней не создаются аноксические условия. Такие условия складываются под пологом лесов некоторых типов и в глубоких тенистых ущельях. Из-за постоянно высокой влажности воздуха транс-пирация у гигрофитов ослаблена, что сильно осложняет транспорт воды по растению и соответственно поглощение корнями минеральных веществ из почвы и снабжение ими надземных частей растения. Поэтому листья гигрофитов устроены так, чтобы максимально усилить транспирацию (рис. 225). Все они имеют тонкую кутикулу, немногочисленные, но крупные почти постоянно открытые устьица. Листовые пластинки тонкие, у некоторых папоротников однослойные за исключением области жилки. Клетки мезофилла располагаются рыхло, даже в зоне палисадной хлоренхимы. Все это увеличивает удельную поверхность, с которой испаряется вода. Устьица располагаются в абаксиальной эпидерме на уровне поверхности листа или вынесены на довольно высоких выступах. Благодаря этому они оказываются за пределами стабильного воздуха, «прилипшего» к поверхности растения, в котором отсутствие турбулентности создает сопротивление диффузии водяного пара через устьица. Усилению транспирации служат также не отмирающие трихомы. Кроме того, гигрофиты рекретируют воду через гидатоды. Несмотря на перечисленные особенности, транспорт воды по телу гигрофитов незначителен, и у многих из них ксилема и корневая система развиты даже слабее, чем у гелофитов. Поскольку гигрофиты не испытывают водного стресса, то основную механическую роль у них играет гидроскелет - тургесцентная основная паренхима, хлоренхима и колленхима, а склеренхима обычно отсутствует. Мезофиты обитают в наиболее распространенных на Земле биотопах со средним значением влажности. Они очень редко и ненадолго оказываются в условиях избытка воды, а регулярный суточный дефицит воды непродолжителен и ненапряжен. Данная экологическая группа объединяет большинство видов наземных растений, вследствие чего все «типичные» черты строения вегетативных органов высших растений - это черты строения органов мезофитов. Ксерофиты - это растения, приспособленные существовать при длительном сезонном дефиците воды в период вегетации. На основании существенных габитуальных и физиологических различий ксерофиты подразделяют на суккуленты и склерофиты, которые реализуют разные стратегии переживания водного дефицита. Суккуленты (рис. 226) приспособлены запасать большое количество воды в короткие влажные периоды и экономно расходовать ее во время засухи. Поэтому у них развита обильная водозапасающая паренхима, вследствие чего их листья (листовые суккуленты) или стебли (стеблевые суккуленты) толстые, сочные и мясистые.
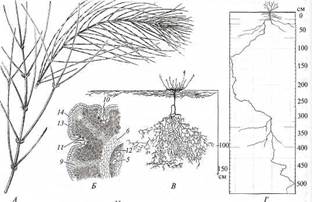
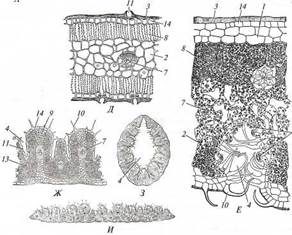
Многие суккуленты имеют поверхностную корневую систему с мощными горизонтальными скелетными корнями, занимающую большую площадь. После дождей на них развиваются сосущие корни, существующие всего 2 — 3 недели, пока в субстрате сохраняется влага. Такая конструкция корневой системы позволяет быстро поглощать довольно скудную влагу осадков, неглубоко промачивающих почву. Ряд видов суккулентов, населяющих туманные пустыни, поглощает конденсат воды с помощью специализированных трихом на листьях и стеблях. Некоторые из них во взрослом состоянии даже не имеют корней. Склерофиты (рис. 227) также содержат запас воды, но в основном в склеренхиме, которая занимает в теле растения большой относительный объем, хотя абсолютный объем ее сравнительно невелик. Поэтому склерофитам свойственны «тощие» жесткие побеги. Главное средство борьбы этих растений с водным дефицитом - создаваемая водоносной склеренхимой высокая сосущая сила, достигающая 60 — 80 (до ПО) атм. Благодаря ей растения могут извлекать воду во время засухи из очень сухого субстрата и прочно удерживать ее в своем теле. Соответственно этому многие виды имеют глубокую корневую систему, проникающую в почву на несколько метров в более насыщенные влагой горизонты вплоть до достижения зеркала грунтовых вод. У некоторых видов корневая система комбинированного типа - с поверхностными и глубоко проникающими в грунт корнями, позволяющая эффективно использовать как влагу глубоких слоев субстрата, так и воду редких дождей, неглубоко промачивающих почву. Общая черта всех ксерофитов - морфолого-анатомические приспособления для снижения транспирации. У многих склерофитов и стеблевых суккулентов листья мелкие, нередко эфемерные, развивающиеся только в короткие влажные сезоны, или редуцированные до небольших чешуек (афилльные виды) либо преобразованные в колючки. Функцию фотосинтеза у них выполняют стебли, имеющие значительно меньшую, чем листья, удельную поверхность и более экономно расходующие воду. В кортексе таких стеблей развиты мощные массивы хлоренхимы из палисадных клеток, перпендикулярных поверхности. Некоторые афилльные склерофиты имеют диморфные побеги: многолетние скелетные более или менее типичного строения и односезонные ассимилирующие со слабым вторичным утолщением, сплошной зоной субэпи-дермальной хлоренхимы и снабжающей ее особой системой кортикальных пучков. Часть склерофитов имеет хорошо развитые длительно функционирующие листья, но они узкие (снижение удельной поверхности) и содержат много склеренхимы в мезофилле (усиление водоудерживающей способности тканей листа). Листья хорошо развиты у листовых суккулентов. Многие ксерофиты густо опушены. Опушение из отмерших, часто ветвистых волосков стабилизирует прилегающий к поверхности растения слой воздуха, в котором не возникают турбулентные движения. Поэтому газы, в том числе и водяной пар, диффундируют в нем значительно медленнее, что снижает интенсивность транспирации через устьица. У некоторых видов опушение сложено водоносными пузыревидными трихомами, смыкающимися друг с другом в почти непрерывный покровный слой над основными клетками эпидермы (см. рис. 226, Г, Д). Устьица этих растений открываются в узкие щелевидные пространства между пузыревидными трихомами. Такие пространства создают высокое диффузионное сопротивление для водяного пара, вследствие чего транспирация растения снижена. Многие суккуленты не имеют опушения, но на поверхности кутикулы у них находится толстый слой воска.
Эпидерма ксерофитов покрыта толстой кутикулой (см. рис. 226, Ж, 227, Д, Е). Основные клетки эпидермы имеют толстую или очень толстую кутинизированную наружную стенку. Устьица «погруженные», т. е. их замыкающие клетки находятся ниже уровня основных клеток. Поэтому апертура устьица располагается на дне ямки, в которой воздух стабилизирован и потому создает повышенное сопротивление диффузии водяного пара через апертуру во внешнюю среду. У многих ксерофитов вокруг устьица имеется валик из кутикулы или выростов основных клеток, который увеличивает глубину ямки и тем самым сопротивление диффузии пара во внешнюю среду (см. рис. 226, Ж, 227, Д). Еще более высокое сопротивление транспирационной потере воды создает воздух в криптах - углублениях на поверхности растения. Устьица располагаются в эпидерме, выстилающей крипты (см. рис. 227, Е, Ж). Между криптами устьиц нет. В криптах развито опушение, еще более стабилизирующее воздух в них. Все это настолько эффективно снижает диффузию газов через устьица, что у некоторых видов устьица в криптах располагаются на выступах эпидермы для облегчения газообмена с внешней средой. В этом проявляются противоречивые тенденции эволюции структур растений, реализующиеся в их компромиссно адаптивной организации. Суккулентам свойственны низкий устьичный индекс, т.е. отношение числа устьиц к числу основных клеток эпидермы, и малое число устьиц на единицу площади органа. У склерофитов, как правило, устьичный индекс такой же, как у мезофитов, но вследствие мелкоклеточности эпидермы число устьиц на единицу площади значительно выше. Однако из-за мелких размеров устьиц удельная суммарная площадь апертур у ксерофитов лишь ненамного превосходит удельную суммарную площадь апертур у мезофитов. Поэтому для суккулентов характерна постоянно низкая транспирация, а склерофиты при полном обеспечении водой транспирируют так же или даже более интенсивно, чем мезофиты, но в условиях водного дефицита они резко снижают транспи-рацию благодаря структурным особенностям и расположению устьиц, а также способности водоносной склеренхимы прочно удерживать воду. Дополнительными возможностями снижать транспирацию обладают склероморфные злаки. Они имеют ребристые с адаксиальной стороны листья (см. рис. 227, Ж - И). Устьица развиты в адаксиальной эпидерме по краям ребер и таким образом открываются в узкие ложбинки-крипты между ребрами. При достатке воды пластинка плоская; в противном случае листовая пластинка сворачивается трубочкой на адаксиальную сторону. В результате вся адаксиальная сторона становится дном временной крипты. В абаксиальной эпидерме устьиц нет, а подстилает ее мощный слой склеренхимы. Вследствие этого транс-гшрация через абаксиальную эпидерму пренебрежимо мала. Изменения протяженности адаксиальной эпидермы и прилегающих к ней тканей мезофилла при сворачивании и разворачивании листовой пластинки обеспечивают варьирование размеров особых крупных сильно вакуолизиро-ванных моторных клеток, располагающихся продольными рядами в адаксиальной эпидерме. Раньше движения пластинки листа считали следствием изменения тургора моторных клеток. Однако оказалось, что они обусловлены различным напряжением в склеренхиме в зависимости от обводненности ее клеток. Эти напряжения вызывают определенные искривления массива склеренхимы и, соответственно, сворачивание или разворачивание пластинки листа. Психрофиты - растения холодных постоянно влажных биотопов, населяющие территории в высоких широтах и высокогорья, где величина осадков превышает величину испарения. По морфолого-анатомическим признакам они сходны с гигро- или мезофитами, но при этом обладают свойствами микро-термных растений. Краофиты также представляют группу микротермных видов, но приспособленных к сухим биотопам и очень сухим горным пустыням и потому имеют ксероморфную организацию. Для растений этой группы в целом характерен медленный рост, для многих — обильное ветвление, вследствие чего возникают распластанные по субстрату плотные дернины или низкие подушковидные кустики. Суккулентные растения не выдерживают глубокого промораживания тканей из-за высокой обводненности. Поэтому большинство криофитов — склерофиты, а суккулентные криофиты встречаются только в тропических горных пустынях, где регулярны заморозки, но не бывает сильных морозов.
|
||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-08-12; просмотров: 2131; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.214 (0.017 с.) |