Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Экологические группы растений по отношению к субстратуСодержание книги
Похожие статьи вашей тематики
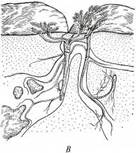
Поиск на нашем сайте Литофиты, или петрофиты, населяют трещины скал и каменистые осыпи (рис. 229, В). Специфика этих биотопов - быстрое просачивание воды по щелям и малое количество мелкозема, удерживающего воду и содержащего минеральные вещества в доступной растению форме. Поэтому растения развивают мощную корневую систему, глубоко проникающую в трещины и щели субстрата за водой и минеральными соединениями. Корневые выделения литофитов содержат повышенное количество кислот, растворяющих породу и способствующих извлечению из нее необходимых веществ. Скудость доступных растению ресурсов и необходимость развивать мощную корневую систему приводят к тому, что организм экономит на развитии побеговой системы. Поэтому побеговая система литофитов непропорционально мала сравнительно с таковой у растений других экологических групп. Так как вода быстро просачивается в недоступные растению горизонты, то литофиты существуют
в условиях устойчивого водного дефицита и имеют небольшие или мелкие густоолушенные кожистые листья для уменьшения транспирации. У обитателей каменистых осыпей развивается очень толстая феллема, уменьшающая травматизм при подвижках камней. Псаммофиты - специализированные обитатели подвижных или слабо закрепленных незадернованных песков (см. рис. 229, А, Б). Вода легко просачивается в глубь песчаного субстрата, унося с собой растворимые минеральные соединения и оставляя бесплодный кварцит. Поэтому псаммофитам присуща мощная корневая система, глубоко проникающая в субстрат или занимающая большую площадь, и ксероморфная, а иногда суккулентная организация побегов. Кроме того, растения на подвижных песках испытывают периодическое засыпание песком или оголение корневой системы из-за выдувания песка. Псаммофиты легко переносят погребение песком благодаря быстро растущим сквозь наметенный песок побегам, развивающим систему придаточных корней на оптимальной глубине. При этом часть тела, оказавшаяся слишком глубоко, отмирает, и растение как бы всплывает в песчаном бугре. Оголенные после выдувания песка корни не отмирают, так как защищены мощным покровом. У двудольных растений он представлен толстой феллемой. Не имеющие феллогена однодольные растения выделяют корнями обильную слизь, склеивающую песчинки, после высыхания которой вокруг корня образуется защитный чехол из сцементированного песка, по толщине в несколько раз превосходящий сам корень. Двудольные псаммофиты способны давать корневые отпрыски на приземных частях оголенных корней, благодаря чему растение опускается вслед за понизившимся уровнем субстрата. Кальцефилы населяют биотопы с щелочной реакцией субстрата и повышенным содержанием в нем СаСО3. Адаптация к таким условиям проявляется на уровне метаболических реакций. Растения некоторых видов рекретируют карбонат кальция через гидатоды. Выраженную структурную специфику проявляют только кальцефилы, обитающие на обнажениях известняка, мела или мергеля. Обнажения известняка представлены скалами и каменистыми осыпями, и обитающие на них кальцефилы, естественно, имеют признаки литофитов. Мергели и мела не только очень легко пропускают воду в недоступные растению горизонты, но и хорошо отражают свет, сильно подсвечивая растения снизу. Поэтому обитающие на таких субстратах кальцефилы проявляют признаки как ксероморфной, так и гелиоморфной организации. В частности, для них характерны изолатеральные листья с мощной многослойной палисадной хлоренхимой с обеих сторон листовой пластинки и тонкой прослойкой губчатой хлоренхимы между зонами палисадной ткани (см. рис. 228, Д). У многих видов кальцефилов на побегах развито густое серебристое опушение, снижающее транспирацию и интенсивно отражающее падающий на растение свет. Нитрофилы - растения биотопов, почвы которых обогащены азотом в легкодоступных растениям формах, главным образом в виде соединений аммония. Эти виды, по-видимому, нуждаются в повышенном количестве азота или не способны поглощать его столь же эффективно, как растения других экологических групп, с которыми они не выдерживают конкуренцию в местах с обычным фоновым содержанием азотных соединений в почве. Напротив, на почвах с повышенным содержанием азота они конкурентно подавляют растения других экологических групп. Более того, нитрофилы хорошо развиваются и при таких высоких концентрациях азотных соединений, которые действуют угнетающе на другие растения. Адаптации нитрофилов к среде обитания проявляются на уровне метаболических реакций и никак не отражаются на их морфолого-анатомической организации. Оксилофиты включают виды, приуроченные к субстратам, бедным минеральными солями: олиготрофным верховым и переходным болотам, торфяникам, задернованным пескам (рис. 230). Они имеют мелкие плотные кожистые листья, покрытые толстой кутикулой. Основные клетки эпидермы - с толстыми кутинизированными наружными стенками. Листовые пластинки в большей или меньшей степени завернуты на абаксиальную сторону, на которой находятся устьица и трихомы. Благодаря этому устьица оказываются изолированными от подвижного воздуха. Все перечисленные особенности сближают оксилофиты с ксерофитами. Однако в отличие от ксерофитов в листьях окси-лофитов развиты крупные воздухоносные полости. Мелкоклеточная хлоренхима дифференцирована на палисадную и губчатую. В листе обычно имеются тяжи склеренхимы, армирующие проводящие пучки и край листовой пластинки.
Ксероморфизм листьев оксилофитов объясняли «физиологической сухостью» мест их обитания, которую трактовали как следствие либо недоступности воды, удерживаемой в гиалиновых клетках сфагнума, либо низкой теплопроводностью сфагнума и торфа, обусловливающей низкую температуру субстрата, что не позволяет мезотермным оксилофитам извлекать из него воду. В последнее время ксероморфизм оксилофитов объясняют малой доступностью минеральных веществ. Из-за этого растения не имеют ресурсов для ежегодной смены листьев и используют листья 2-3 вегетационных сезона. Ксероморфная структура листа защищает его от высыхания в периоды анабиоза, когда испарение продолжается, а транспорт воды по растению прекращается. (Этим же обусловлено ксероморфное строение хвои у хвойных мезофитов (ель, пихта и др.). Кроме того, в листьях оксилофитов клеточные стенки занимают значительный удельный объем вследствие мелкоклеточности листа и обилия в нем склеренхимы. Это снижает пищевую ценность такого листа для фитофагов. Этому же способствуют многочисленные железистые трихомы, вырабатывающие ядовитый секрет. В результате фитофаги почти не повреждают листья оксилофитов, и растение не расходует ресурсы на восстановление листвы. Оксилофиты, обитающие на сфагновых болотах, постепенно погребаются нарастающим сфагнумом. Поэтому они нарастают согласованно с ростом сфагнума и многие, подобно псаммофитам, развивают многочисленные придаточные корни. Галофиты представлены обитателями субстратов с повышенным содержанием хлоридов и/или сульфатов щелочных и щелочноземельных металлов(рис. 231). Их общая черта — высокая сосущая сила. У одних видов она возникает благодаря высокой концентрации солей и/или низкомолекулярных органических веществ в клеточном соке, а у других — вследствие упругой деформации стенок склерифицированных тканей. Другие признаки адаптации галофитов к засолению разнообразны, на основании чего эту группу подразделяют на три подгруппы.
Криптогалофиты, или солевыделяющие галофиты, рекретируют поглощаемые соли. Для этого у них есть специализированные солевые железки или трихомы с крупной сильно вакуолизированной терминальной клеткой (см. рис. 231, В, Г). Железки активно выделяют на поверхность концентрированный рассол, который быстро высыхает, после чего кристаллы соли постепенно опадают с растения. Трихомы накапливают соли в вакуоле терминальной клетки, после чего отмирают и опадают с растения. В обоих случаях растение освобождается от избытка солей и потому иных признаков галоморфнои организации не проявляет. По общей организации криптогалофиты сходны со скле-рофитами или мезофитами. Эвгалофиты, или соленакаплавающие галофиты, выделяют избыточные соли в вакуоли клеток специализированной паренхимы, состоящей из крупных сильно вакуолизиро-ванных клеток. Большой объем этой ткани придает мясистость органам эвгалофитов и сходство с суккулентами (рис. 231, А, Б). Однако накопление большого количества воды в специализированной паренхиме эвгалофитов связано с отложением в ней солей, а не с запасанием воды, так как эти растения часто живут на влажном или даже затопленном грунте. Поэтому некорректно называть эту паренхиму водозапасающей, как это нередко делают. Она представляет собой не запасающую, а рекреторную ткань. В отличие от настоящих суккулентов эвгалофиты не экономят воду и имеют тонкую кутикулу и довольно тонкие наружные стенки основных клеток эпидермы. Кроме того, в связи с очень высоким осмотическим давлением (до нескольких сотен атмосфер) клеточного сока ксилема эвгалофитов сильно склерифицирована, что типичным суккулентам не свойственно. Таким образом, эвгалофиты следует относить к галосуккулентам, структурно напоминающим суккуленты, но принципиально отличающимся от них физиологией. Гликогалофиты, или солеисключающие галофиты, благодаря особым свойствам плазмалеммы клеток ризодермы поглощают почти пресную воду из глубоких слабозасоленных горизонтов почвы. Соответственно в организации остальных тканей и органов гликогалофитов адаптации к засолению не выражены. Эпифиты приспособлены к существованию на поверхности тела других растений. Сосудистые высшие растения обитают на стволе и ветвях деревьев (рис. 232). Разные виды мохообразных заселяют стволы, ветви и листья. Для поверхности растения как субстрата характерно быстрое етекание воды и крайняя скудость элементов минерального питания. Поэтому сосудистые эпифиты обладают суккулентной организацией, а корни многих видов (орхидей) покрыты веламеном, обеспечивающим быстрое впитывание дождевой воды и конденсата. Некоторые эпифиты впитывают влагу с помощью особых трихом, густо покрывающих стебли и листья. Эпифитные мохообразные пойкилогид-ричны и в промежутках между дождями впадают в анабиоз, высыхая до воздушно-сухого состояния. Многие эпифиты задерживают воду в розетке листьев, в асцидиатных листьях (см. рис. 77, Б) или в «кармане» между веткой растения-опоры и прижатыми к ней специализированными неассимилирую-щими листьями (рис. 232, Б). В таких вместилищах со временем накапливается некоторое количество опада и образуется подобие почвы или, точнее, ила. В него могут прорастать придаточные корни, что улучшает питание эпифита.
|
|||||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-08-12; просмотров: 3261; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.214 (0.013 с.) |