Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Лизогения и её биологическое значениеСодержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
Бактериофаги Бактериофаги (фаги) – это вирусы, поражающие клетки бактерий. Они не имеют клеточной структуры, неспособны сами синтезировать нуклеиновые кислоты и белки, поэтому являются облигатными внутриклеточными паразитами. Вирионы фагов состоят из головки, содержащей нуклеиновую кислоту вируса, и отростка. Нуклеокапсид головки фага имеет кубический тип симметрии, а отросток – спиральный тип, т. е. бактериофаги имеют смешанный тип симметрии. Фаги могут существовать в двух формах: 1) внутриклеточной (это профаг, чистая ДНК); 2) внеклеточной (это вирион). Фаги, как и другие вирусы, обладают антигенными свойствами и содержат группоспецифические и типоспецифические антигены. Различают два типа взаимодействия фага с клеткой: 1) литический (продуктивная вирусная инфекция). Это тип взаимодействия, при котором происходит репродукция вируса в бактериальной клетке. Она при этом погибает. Вначале происходит адсорбция фагов на клеточной стенке. Затем следует фаза проникновения. В месте адсорбции фага действует лизоцим, и за счет сократительных белков хвостовой части в клетку впрыскивается нуклеиновая кислота фага. Далее следует средний период, в течение которого подавляется синтез клеточных компонентов и осуществляется дисконъюнктивный способ репродукции фага. При этом в области нуклеоида синтезируется нуклеиновая кислота фага, а затем на рибосомах осуществляется синтез белка. Фаги, обладающие литическим типом взаимодействия, называют вирулентными. В заключительный период в результате самосборки белки укладываются вокруг нуклеиновой кислоты и образуются новые частицы фагов. Они выходят из клетки, разрывая ее клеточную стенку, т. е. происходит лизис бактерии; 2) лизогенный. Это умеренные фаги. При проникновении нуклеиновой кислоты в клетку идет интеграция ее в геном клетки, наблюдается длительное сожительство фага с клеткой без ее гибели. При изменении внешних условий могут происходить выход фага из интегрированной формы и развитие продуктивной вирусной инфекции. Клетка, содержащая профаг в геноме, называется лизогенной и отличается от исходной наличием дополнительной генетической информации за счет генов профага. Это явление лизогенной конверсии. По признаку специфичности выделяют: 1) поливалентные фаги (лизируют культуры одного семейства или рода бактерий); 2) моновалентные (лизируют культуры только одного вида бактерий); 3) типовые (способны вызывать лизис только определенных типов (вариантов) бактериальной культуры внутри вида бактерий). Фаги могут применяться в качестве диагностических препаратов для установления рода и вида бактерий, выделенных в ходе бактериологического исследования. Однако чаще их применяют для лечения и профилактики некоторых инфекционных заболеваний. 1 Структура и принципы классификации бактериофагов. Бактериофаги (пожиратель) — вирусы бактерий, обладающие способностью специфически проникать в бактериальные клетки, репродуцироваться в них и вызывать их растворение (лизис). Морфология. Имеет двунитчатую ДНК с сокращаюмщимся отростком. Форма зависит от типа симметрии капсомера. Если кубический–то форма многогранника, если по спирали то форма палочковидная. Имеют форму головастика или сперматозоида, кубическую и нитевидную формы. Имеющие форму сперматозоида состоят из вытянутой икосаэдрической головки и хвостового отростка. Внутри хвостового отростка имеется полый цилиндрический стержень, сообщающийся отверстием с головкой, снаружи — чехол, способный к сокращению наподобие мышцы. Хвостовой отросток заканчивается шестиугольной базальной пластинкой с короткими шипами, от которых отходят нитевидные структуры — фибриллы. Химический состав. нуклеиновой кислоты (ДНК или РНК) и белка. У фагов, имеющих форму сперматозоида, двунитчатая ДНК упакована в виде спирали внутри головки. Белки входят в состав оболочки (капсида), окружающей нуклеиновую кислоту, и во все структурные элементы хвостового отростка. Обнаружены внутренние (геномные) белки, связанные с нуклеиновой кислотой, и белки-ферменты (лизоцим, АТФ-аза), участвующие во взаимодействии фага с клеткой. Резистентность. Ряд дезинфицирующих веществ (фенол, этиловый спирт, эфир и хлороформ) не оказывают существенного влияния на фаги. Высокочувствительны к формалину и кислотам. Инактивация наступает при температуре 65—70 °С, сохраняются при высушивании в запаянных ампулах, замораживании при температуре —185 °С в глицерине. По механизму взаимодействия различают вирулентные и умеренные фаги. При попадании в бактерию по своему взаимодействию могут быть вирулентными,т.е. попав в клеткуидет репродукция вируса и лизис бактерии. Некоторые фаги попав в бактерию не репродуцируются в ней, не вызывая ее лизиса, а их ДНК включается в бактериальную хромосому это называется профаг. А сам фаг назыв. умеренным. А это явление (биологический симбиоз микробной клетки с умеренным фогом (профагом) назыв. Лизогения, а культура бактерий, содержещий профаг, назыв. лизогенной. Лизогения это когда под действием внеш. факторов профаг выходит из интегрированного состояния и превращается в вирулентный,т.е. идет репродукция фаговых частиц и лизис. Фаг включается в ДНК (нуклеотид) и может взять кусок ДНК и передать другому – фаговая конверсия ( мощный фактор изменчивости микроорганизмов). По специфичности действия различают поливалентные фаги, способные взаимодействовать с родственными видами бактерий, моновалентные фаги, взаимодействующие с бактериями определенного вида, и типовые фаги, взаимодействующие с отдельными вариантами (типами) данного вида бактерий. 2 Механизм взаимодействия бактериофагов с клеткой. По механизму взаимодействия различают вирулентные и умеренные фаги. Вирулентные ф аги, проникнув в бактериальную клетку, автономно репродуцируются в ней и вызывают лизис бактерий. Эти фаги адсорбируются на поверхности бактериальной клетки с помощью фибрилл хвостового отростка. В результате активации фагового фермента АТФазы происходит сокращение чехла хвостового отростка и внедрение стержня в клетку. В процессе “прокалывания” клеточной стенки бактерии принимает участие фермент лизоцим, находящийся на конце хвостового отростка. Вслед за этим ДНК фага, содержащаяся в головке, проходит через полость хвостового стержня и активно впрыскивается в цитоплазму клетки. Остальные структурные элементы фага (капсид и отросток) остаются вне клетки. После биосинтеза фаговых компонентов и их самосборки в бактериальной клетке накапливается до 200 новых фаговых частиц. Под действием фагового лизоцима и внутриклеточного осмотического давления происходит разрушение клеточной стенки, выход фагового потомства в окружающую среду и лизис бактерии. Один литический цикл (от момента адсорбции фагов до их выхода из клетки) продолжается 30—40 мин. По специфичности действия различают поливалентные фаги, способные взаимодействовать с родственными видами бактерий, моновалентные фаги, взаимодействующие с бактериями определенного вида, и типовые фаги, взаимодействующие с отдельными вариантами (типами) данного вида бактерий. Умеренные фаги лизируют не все клетки в популяции, с частью из них они вступают в симбиоз, в результате чего ДНК фага встраивается в хромосому бактерии. В таком случае геномом фага называют профаг. Профаг, не вызывая ее лизиса, передается по наследству от клетки к клетке. Биологическое явление симбиоза микробной клетки с умеренным фагом (профагом) называется лизогенией, а культура бактерий, содержащая профаг, получила название лизогенной. Это отражает способность профага самопроизвольно или под действием ряда физических и химических факторов исключаться из хромосомы клетки и переходить в цитоплазму, т. е. вести себя как вирулентный фаг, лизирующий бактерии. Лизогенные культуры невосприимчивы к повторному заражению близкородственным фагом и приобретают дополнительные свойства, которые находятся под контролем генов профага. Изменение свойств микроорганизмов под влиянием профага получило название фаговой конверсии и касается различных их свойств: культуральных, биохимических, токсигенных, антигенных, чувствительности к антибиотикам и др. Кроме того, переходя из интегрированного состояния в вирулентную форму, умеренный фаг может захватить часть хромосомы клетки и при лизисе последней переносит эту часть хромосомы в другую клетку. Если микробная клетка станет лизогенной, она приобретает новые свойства. Таким образом, умеренные фаги являются мощным фактором изменчивости микроорганизмов Практическое использование бактериофагов в медицине и микробиологии. Применение фагов основано наихстрогой специфичности действия. Фаги используют в диагностике инфекционных болезней: 1) с помощью известных (диагностических) фагов проводят идентификацию выделенных культур микроорганизмов. Вследствие высокой специфичности фагов можно определить вид возбудителя или варианты (типы) внутри вида. Фаготипирование имеет большое эпидемиологическое значение, так как позволяет установить источник и пути распространения инфекции; 2) с помощью тест-культуры можно определить неизвестный фаг в исследуемом материале, что указывает на присутствие в нем соответствующих возбудителей. Фаги применяют для лечения и профилактики инфекционных болезней. Производят брюшнотифозный, дизентерийный, синегнойный, стафилококковый фаги и комбинированные препараты. Способы введения в организм: местно, энтерально или парентерально. Умеренные фаги используют в генетической инженерии и биотехнологии в качестве векторов для получения рекомбинантных ДНК. Дефектный фаг учавствует в трансдукции кот. дает: 1) резистентность к лекарст.препаратам.; 2) образование капсулы; 3) синтез ферментов ращепляющих углеводы. 3-и вида трансдукции: 1) генерализованная. Когда умеренный фаг встраивается в донора в любом месте хромосомы и забирает разные участки и св-ва; 2) спецефическая есть фаг он встраивается в одном месте и переносит всегда ген-способность синтезировать фермент галактоза; 3) абортивная форма. Когда при делении кл. фаговая ДНК передаётся только одной клетки и признак теряется, т.е. нет репликации данного участка. Изменение свойств микроорганизмов под влиянием профага получило название фаговой конверсии и касается различных их свойств: культуральных, биохимических, токсигенных, антигенных, чувствительности к антибиотикам и др. Умеренные фаги могут нанести вред микробиологическому производству. Так, если микроорганизмы, используемые в качестве продуцентов вакцин, антибиотиков и других биологических веществ, оказываются лизогенными, существует опасность перехода умеренного фага в вирулентную форму, что неминуемо приведет к лизису производственного штамма. Бактериофаги и актинофаги Во второй половине прошлого столетия учеными-микробиологами было открыто большинство бактерий, вызывающих заболевания человека, животных и растений. Были разработаны методы выделения этих бактерий и размножения их на искусственных питательных средах. В то же время микробиологи обратили внимание на ряд инфекционных заболеваний человека и животных (оспа, бешенство, корь, краснуха и др.), а также растений, при которых бактериальные методы выяснения их возбудителей оказались непригодными. Выходом из тупика, в котором очутилась микробиологическая наука, послужило открытие, сделанное русским ученым Д. И. Ивановским. В 1887 г. он, будучи еще студентом Петербургского университета, поехал на Украину и в Молдавию, где изучал причины мозаичной болезни табака, которая причиняла большой ущерб табачным плантациям южных районов. Свои исследования он проводил в течение нескольких лет и после окончания университета. В результате тщательного изучения этого заболевания Д. И. Ивановский показал, что оно вызывается специфическим возбудителем, состоящим из мельчайших частиц, или корпускул, которые по своим размерам значительно меньше бактерий. Листья табака, пораженного мозаикой, он растирал в ступке и затем полученную массу фильтровал через специальные бактериальные фильтры, не пропускающие бактерий. Полученная после фильтрации прозрачная жидкость не содержала видимых в обычном микроскопе частиц. Но при нанесении ее на царапины, сделанные на поверхности здоровых листьев табака, растение заболевало мозаичной болезнью. Этот прием можно было повторять много раз. Результаты исследований Д. И. Ивановского были опубликованы в 1892 г. в книге «О двух болезнях табака». В этой работе впервые в истории микробиологии было показано, что в природе существует особый мир мельчайших возбудителей инфекционных заболеваний. Эти возбудители проходят через бактериальные фильтры, невидимы в обычном микроскопе, не растут на тех средах, которые применяются для выращивания бактерий, и способны размножаться только в организме человека, животных и растений. Эти мельчайшие организмы получили название фильтрующихся вирусов или вирусов. Блестящее открытие Д. И. Ивановского ознаменовало новую эпоху в развитии микробиологии и заложило основы новой науки — вирусологии. Открытие первого вируса — возбудителя мозаики табака — было толчком к проведению широких исследований в области вирусологии. Пользуясь предложенным Д. И. Ивановским методом, ученые стали открывать один за другим вирусы, вызывающие различные заболевания человека, животных и растений. В конце XIX — начале XX в. стало известно, что в природе существуют также вирусы, которые поражают бактерии и при этом вызывают распад (лизис) их клеток. Эти вирусы получили название бактериофагов — «пожирателей бактерий». Явление лизиса, или распада, микробной клетки было известно давно. Оно может быть вызвано различными физическими и химическими факторами. В ряде случаев распад может происходить на определенном этапе развития микробной клетки как нормальный физиологический процесс ее старения. Однако имеется принципиальное различие между распадом клетки, вызванным бактериофагами, и химическими факторами. В первом случае происходит размножение бактериофага, вызывающего лизис, что не происходит при распаде клетки под влиянием физических и химических факторов. Одним из первых, кто наблюдал и детально описал явление лизиса у бактерий, был один из основоположников отечественной медицинской микробиологии — Н. Ф. Гамалея. В 1896 —1898 гг. появились его работы, посвященные изучению явления лизиса у сибиреязвенной палочки. Фактор, вызвавший лизис этой бактерии, он назвал бактериолизином. Для развития исследований в области бактериофагии особое значение имели работы французского ученого Д'Э р е л л я. В 1917 г. он сообщил, что из фекальных масс больных дизентерией ему удалось выделить особый лити-ческий фактор (вирус), способный проходить через бактериальные фильтры, размножаться на дизентерийных бактериях и вызывать при этом их лизис. Для обозначения этого вируса Д'Эрелль впервые предложил название бактериофаг. Кроме названия бактериофаг, или (сокращенно) фаг, в литературе, особенно в более старой, можно встретить также следующие: бактериофагический лизин, феномен Д'Эрелля, феномен Туорта, феномен Д'Эрел-ля — Туорта. Название «феномен Туорта» связано с именем английского микробиолога Туорта, который в 1915 г. описал явление перевиваемого лизиса у стафилококков и высказал предположение о вирусной природе этого явления. Для обозначения фагов (вирусов микроорганизмов), вызывающих лизис актиномицетов, применяется термин актинофаг, микобакте-рий— микофаг, кишечной палочки — колифаг, водорослей — цианофаг и т. д. Вначале много внимания уделялось изучению фагов, активных против патогенных бактерий: дизентерийной, брюшнотифозной, дифтерийной палочек, стафилококков с целью выяснения возможности использования их для лечения и профилактики инфекционных заболеваний. Одновременно много внимания уделялось изучению природы фагов. Однако в последние годы проблема бактериофагии фактически превратилась в самостоятельную область биологии со своими специфическими разделами. Фаги оказались весьма удобной моделью для разрешения ряда важнейших теоретических и практических вопросов общей биологии, генетики, молекулярной биологии, биохимии, а также медицины, ветеринарии и вирусологии. Некоторые фаги отличаются специфичностью, т. е. они способны лизировать только определенные виды и разновидности бактерий. Поэтому такие фаги, названные типовыми, успешно применяются при дифференциации микроорганизмов. Созданы специальные музеи типовых фагов. В результате большого теоретического и практического значения проблемы бактериофагии за последние 10—20 лет фаги изучались весьма интенсивно и всесторонне. Морфология фагов Применение современных электронных микроскопов, а также усовершенствование методов приготовления препаратов для электронной микроскопии позволили более детально изучить тонкую структуру фагов. Оказалось, что она весьма разнообразна и у многих фагов более сложна, чем структура вирусов растений и ряда вирусов человека и животных.
Разные фаги отличаются друг от друга не только по форме, величине и сложности своей организации, но и по химическому составу. Оказалось, что фаги, лизирующие микроорганизмы различных групп, могут быть вполне идентичными по своей морфологии. В то же время фаги, активные против одной и той же культуры, могут резко различаться по своей структуре. Так, например, среди фагов, способных лизировать разные штаммы кишечной палочки, выявлены все известные морфологические типы фагов.
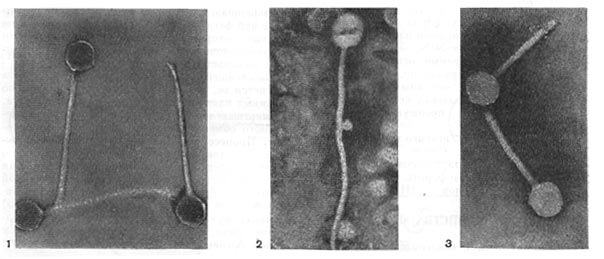
Частицы (или вирионы) большинства известных фагов имеют форму сперматозоида. Они состоят из головки (или капсида) и отростка. Наряду с этим есть фаги, которые состоят из одной головки, без отростка, и фаги, имеющие форму палочки (палочковидные или нитевидные фаги).
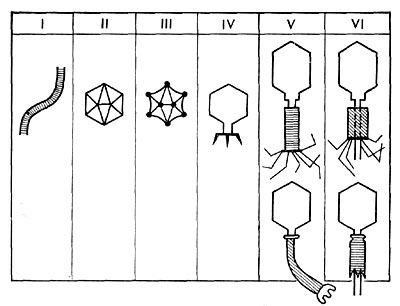
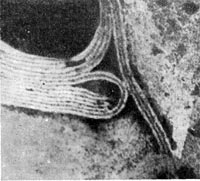
По форме частиц фаги делятся на шесть основных морфологических типов (групп): палочковидные или нитевидные фаги; фаги, состоящие из одной головки, без отростка; фаги, состоящие из головки, на которой имеется несколько небольших выступов; фаги, состоящие из головки и весьма короткого отростка; фаги, имеющие головку и длинный отросток, чехол которого не может сокращаться; фаги, имеющие головку и длинный отросток, чехол которого может сокращаться. На рисунке 204 показано схематическое изображение частиц всех шести типов фагов. Размеры фагов принято обозначать в милли-микрометрах (1 миллимикрометр — миллионная часть миллиметра) или в ангстремах (10 А = 1 миллимикрометр). Фаги первого морфологического типа — палочковидные или нитевидные — выявлены у кишечной, синегнойной, чудесной палочек и других бактерий. Средние размеры их: длина — от 7000 до 8500 А, ширина — от 50 до 80 А (рис. 205). Эти фаги отличаются от всех остальных не только большой специфичностью, но и рядом других важных свойств (о чем будет сказано дальше).
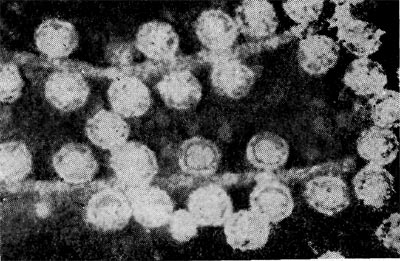
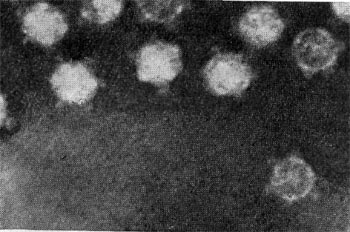
Фаги второго морфологического типа. Частица их состоит из одной головки гексагональной (шестигранной) формы на плоскости. Частицы очень мелкие, средний размер их 230—300 А в диаметре (рис. 206). У фагов третьего морфологического типа форма и размеры головки такие же, как у фагов второго типа, но у их головок имеются обычно несколько очень коротких выступов (рис. 207). Возможно, эти выступы являются аналогами отростков.
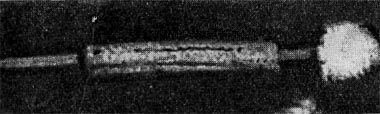
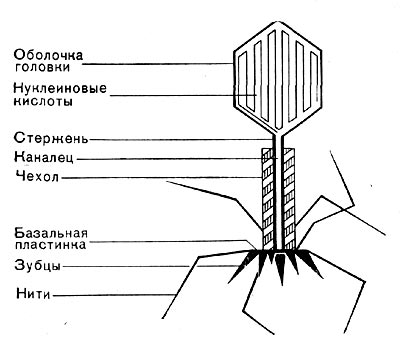
Фаги второго и третьего морфологических типов отличаются постоянством формы и размеров, независимо от того, против каких микроорганизмов они активны. Эти фаги относятся к мелким формам. Фаги четвертого морфологического типа. Частица состоит из головки, размеры которой варьируют от 400 до 640 А в диаметре, и очень короткого отростка (рис. 208). Длина и ширина отростка от 70 до 200 А. Фаги пятого морфологического типа наиболее широко распространены. Головка у частиц гексагональной, формы различных размеров — от 500 до 4250 А в диаметре. Размеры отростка: длина — от 1700 до 5000 А, ширина — от 70 до 120 А (рис. 209). Чехол отростка не способен сокращаться. Фаги шестого морфологического типа также широко распространены. Головка частицы разо-личной формы и размеров — от 600 до 1500 А в диаметре, гексагональная. Размеры отростка: длина — от 800 до 2890 А, ширина — от 140 до 370 А. Важной особенностью фагов этой группы является то, что чехол, окружающий отросток, способен сокращаться, в результате чего становится видимым внутренний стержень отростка (рис. 210). Головки всех фагов состоят из внутреннего содержимого — нуклеиновой кислоты - - и окружены белковой оболочкой. Отросток фагов весьма сложен. Он обычно состоит из следующих структур: наружного чехла (или оболочки), внутреннего стержня с канальцем, базальной пластинки, оканчивающейся выступами (типа шипов) и нитевидными структурами. Чехол отростка состоит из субъединиц белковой природы, собранных в спираль. В результате этого он приобретает вид гофрированной трубки. В верхней части отростка многих фагов имеется образование, которое называется воротничком. На рисунке 211 схематически изображена тонкая структура фаговой частицы. Химический состав фагов Изучение химического состава фагов стало возможно лишь тогда, когда были усовершенствованы методы получения в больших количествах очищенных препаратов фага. В настоящее время изучен химический состав фагов, принадлежащих к разным морфологическим типам и поражающих микроорганизмы почти всех систематических групп. Основными компонентами фагов являются белки и нуклеиновые кислоты. Важно отметить, что фаги, как и другие вирусы, содержат только один тип нуклеиновой кислоты — дез-оксирибонуклеиновую (ДНК) или рибонуклеиновую (РНК). Этим свойством вирусы отличаются от микроорганизмов, содержащих в клетках оба типа нуклеиновых кислот.
Нуклеиновая кислота находится в головке. Внутри головки фагов обнаружено также небольшое количество белка (около 3%). Таким образом, по химическому составу фаги являются нуклеопротеидами. В зависимости от типа своей нуклеиновой кислоты фаги делятся на ДНК-овые и РНК-овые. Количество белка и нуклеиновой кислоты у разных фагов разное. У некоторых фагов содержание их почти одинаковое и каждый из этих компонентов составляет около 50%. У других фагов соотношение между этими основными компонентами может быть различно.
Кроме указанных основных компонентов, фаги содержат в небольших количествах углеводы и некоторые преимущественно нейтральные жиры.
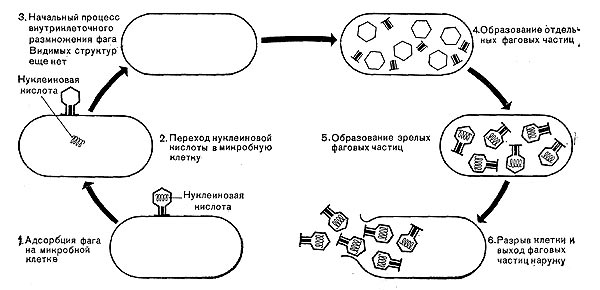
Все известные фаги второго морфологического типа РНК-овые. Среди фагов третьего морфологического типа встречаются как РНК-овые, так и ДНК-овые формы. Фаги остальных морфологических типов — ДНК-овые. Антигенные свойства фагов Известно, что при введении в организм животного подкожно или внутривенно белка, бактериальных клеток, некоторых продуктов жизнедеятельности микроорганизмов и других веществ в крови животного вырабатываются вещества, названные антителами. Вещества, способные вызывать образование антител, называются антигенами. Антитела очень специфичны и способны вступать в реакции только с теми антигенами, которые вызвали их образование. Они или связывают соответствующие антигены, или нейтрализуют их, или осаждают, или растворяют. Оказалось, что все фаги обладают антигенными свойствами. При введении фага в организм животного в сыворотке крови образуются специфические антитела, способные действовать только против данного фага. Такие сыворотки называются антифаговыми. Когда фаг смешивается со специфической антифаговой сывороткой, происходит инактивация фага — фаг теряет способность вызывать лизис чувствительных к нему микробов. Так как каждая антифаговая сыворотка специфична, ее можно успешно применять для идентификации и классификации фагов и очистки микробной культуры от фага. При помощи сыворотки удалось доказать, что белок оболочки фага отличается от белка оболочки отростка и от белка базальной пластинки и ее нитевидных образований, что говорит о сложности структуры фаговой частицы. По антигенным свойствам фаг резко отличается от чувствительных к нему микробов. Размножение фагов Взаимоотношения между фагом и чувствительной к нему клеткой очень сложны и не всегда завершаются лизисом клетки и размножением в ней фага. Мы рассмотрим такую инфекцию клетки, которая заканчивается гибелью клетки и размножением в ней фага. Такая инфекция называется продуктивной. Важнейшей особенностью размножения фага является то, что оно может происходить только в живых клетках, находящихся в стадии роста. В мертвых клетках, а также продуктах клеточного обмена размножение фага не происходит. Процесс размножения фага весьма сложный и состоит из следующих последовательно протекающих этапов (рис. 212): 1) адсорбция фаговой частицы на поверхности микробной клетки; 2) проникновение содержимого головки фаговой частицы (нуклеиновой кислоты) в микробную клетку; 3) внутриклеточное развитие фага, заканчивающееся образованием новых фаговых частиц; 4) лизис клетки и выход из нее новых фагов. Время с момента инфицирования клетки фагом до лизиса клетки называется латентным или скрытым периодом. Продолжительность этого периода различна для разных типов фага, зависит от окружающей температуры, состава среды и других факторов. Латентный период фагов, специфичных для одних бактерий, 15—40 мин, для других — 5 ч и более. У фагов актиномицетов латентный период может быть еще продолжительнее. При низкой температуре латентный период значительно увеличивается.
Из всех этапов размножения фага наиболее изучен первый — адсорбция. Адсорбция фага на клетке — реакция весьма специфичная. В клеточной стенке бактерий имеются особые структуры (рецепторы), к которым могут прикрепиться фаги. Адсорбируются на рецепторах только те фаги, к которым чувствительна клетка. Фаги, имеющие отростки, прикрепляются к микробной стенке свободным концом отростка. Нитевидные фаги, а также фаги, не имеющие отростков, адсорбируются не на микробной стенке, а на нитевидных структурах, окружающих стенку, — фимбриях. Описаны фаги, которые прикрепляются отростком к бактериальным жгутикам. У некоторых фагов процесс адсорбции может осуществляться лишь в том случае, когда в среде имеются определенные вещества — кофакторы: аминокислоты (триптофан, тирозин и др.) или соли (кальциевые, магниевые). На конце фагового отростка имеется особый фермент типа лизоцима. После адсорбции фага под влиянием этого фермента происходит растворение стенки микробной клетки и содержимое головки фага — нуклеиновая кислота — перекачивается в микробную клетку. Этим завершается второй этап процесса размножения фага. Остальные структуры фаговой частицы — оболочка головки, отросток и его субструктуры — внутрь инфицированной фагом клетки не попадают. Их роль заключается в обеспечении сохранности фаговой частицы, находящейся вне клетки, и содействии проникновению фаговой нуклеиновой кислоты в клетку при инфекции.
У нитевидных фагов, в отличие от других видов фагов, внутрь клетки проникает весь белок или его часть. После проникновения нуклеиновой кислоты фага в клетку начинается сложный процесс внутриклеточного размножения фага. Под влиянием нуклеиновой кислоты фага резко изменяется весь обмен микробной клетки. Основные процессы, протекающие в инфицированной клетке, направлены на образование новых фаговых частиц. Вначале формируются отдельно головки и отростки, которые затем объединяются в зрелые фаговые частицы. К этому времени внутри клетки образуется особый литический фермент, который вызывает лизис клетки изнутри. Клетка распадается, и новые зрелые частицы фага выходят наружу.
Количество новых фаговых частиц, образуемых одной клеткой при фаговой инфекции, называют выходом фага или его урожайностью. Выход фага зависит от свойств данного фага и не зависит от клетки-хозяина и ее размеров. Одни фаги отличаются очень низким выходом (5—50 частиц на клетку), у других выход значительно выше (от 1000 до 2500). Особенно высоким выходом отличаются мелкие РНК-овые фаги (свыше 20 000 частиц на клетку). Если большое количество бактериальных клеток смешать с небольшим количеством фаговых частиц, то процесс размножения фагов проходит несколько циклов. Вначале инфицируется часть клеток. Первое потомство фага инфицирует оставшиеся клетки — происходит второй цикл, за ним может следовать третий и т. д., пока не будут лизированы все чувствительные к данному фагу клетки. Среди фагов встречаются такие, размножение которых возможно лишь при наличии в среде определенных кофакторов. Одни из этих веществ, как уже указывалось, необходимы для адсорбции фага; другие — для внутриклеточного размножения фага.
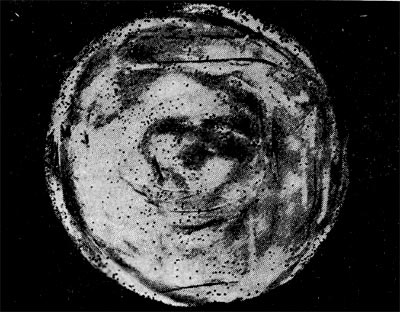
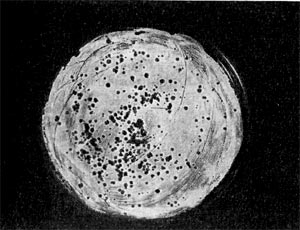
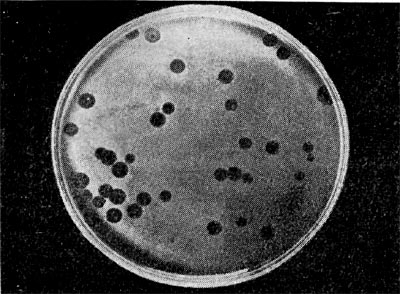
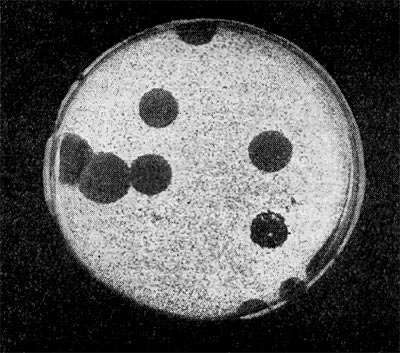
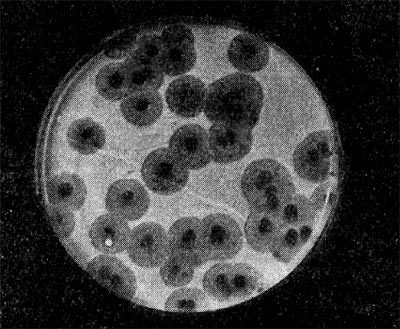
Является ли процесс размножения фага всегда смертельным для инфицированной им клетки? Несомненно, абсолютное большинство фагов вызывают при размножении лизис клетки и ее гибель. Лишь в последнее время было установлено, что при инфицировании клетки нитевидными фагами возможно размножение их без гибели клетки. Если произвести рассев по поверхности ага-ризованной питательной среды в чашках Петри смеси фага и чувствительных к нему микробов и чашки выдержать в термостате, то происходит лизис клеток в результате размножения фага. Если взять большое количество частиц фага, то лизируется большая часть или весь выросший газон культуры. Если количество фаговых частиц таково, что они распределяются только на отдельных участках газона, лизируя в этих местах культуру, то возникает колония фага. Эти колонии фага получили название бляшек, стерильных пятен. Правильнее их называть негативными колониями. Каждая негативная колония состоит из десятков и сотен миллионов фаговых частиц. Размер негативных колоний и их форма зависят в первую очередь от свойств фага, а также от состава среды и культуры микробов. У одних фагов негативные колонии очень мелкие и еле видимы невооруженным глазом, другие достигают 10 мм в диаметре и более. Колонии бывают светлые и четкие, когда лизировалась вся культура, или мутноватые, когда лизировались не все клетки. Вокруг негативных колоний некоторых фагов могут возникнуть различной формы и величины ореолы. На рисунках 213— 217 показаны негативные колонии разных фагов. Морфология негативных колоний служит одним из признаков, которым пользуются при дифференциации фагов. Распространение фагов В настоящее время найдены, фаги, лизирую-щие клетки микроорганизмов, принадлежащих ко всем систематическим группам, как патогенных для человека, животных и растений, так и сапрофитных (непатогенных). До недавнего времени не было ясно, существуют ли фаги против плесневых грибов и дрожжей. В последние годы найдены фаги, активные против грибов родов пенициллов, аспергил-лов и других, а также против некоторых дрожжей. Интересно отметить, что вирус удалось выявить и у тех видов пенициллов, которые применяются в промышленности для получения пенициллина. Не выявлены вирусы, активные против простейших животных, а также истинных спирохет.
В природных условиях фаги встречаются в тех местах, где есть чувствительные к ним бактерии. Чем богаче тот или иной субстрат (почва, вода, выделения человека и животных и т. д.) микроорганизмами, тем в большем количестве в нем встречаются соответствующие фаги. Так, фаги, лизирующие клетки всех видов почвенных микроорганизмов, находятся в почвах. Особенно богаты фагами черноземы и почвы, в которые вносились органические удобрения. Фаги, активные против разных видов кишечной, дизентерийной, тифозной и паратифозной палочек, часто встречаются в содержимом кишечника человека и животных, сточных водах и загрязненных водоемах. Фаги фитопатогенных микроорганизмов успешнее всего выделяются из остатков растений, пораженных этими микробами.
Фаги молочнокислых стрептококков в большом количестве встречаются в молочных продуктах. Итак, те субстраты, на которых развиваются определенные формы микроорганизмов, также благоприятны для существования соответствующих фагов. Трансдукция (перенос) При размножении определенных умеренных фагов на чувствительних к ним культурах фаговая частица захватывает какой-нибудь фрагмент генетического материала данной клетки. При воздействии этим же фагом на другую чувствительную к нему культуру он передает новой культуре захваченный фрагмент. Культура, от которой фаг переносит генетический материал, получила название донора (дающая), а культура, приобретающая генетический материал, — реципиента (получающая). При трансдукции фаг играет роль механического переносчика; лизогенизация клетки не обязательна. Один и тот же фаг может переносить разные свойства. Трансдукция происходит довольно редко: из одного и более миллионов фаговых частиц только одна способна осуществлять трансдук-цию. При помощи трансдукции удавалось перенести от клеток-доноров клеткам-реципиентам различные свойства: токсичность, устойчивость к антибиотикам, способность продуцировать определенные ферменты, антигенные и другие свойства. Природа фагов Несмотря на то что явление бактериофагии интенсивно изучается более пятидесяти лет, на природу фагов нет единой точки зрения и этот вопрос до сих пор остается спорным. В нашей стране и за рубежом в специальной печати и на конференциях неоднократно возникали горячие дискуссии на эту тему. И это не случайно. Вопрос о происхождении фагов, как и других вирусов, имеет большое значение, так как с ним тесно связано решение многих актуальнейших задач современной биологии: происхождение жизни, возможные формы существования живого; существование живых существ, не имеющих клеточной структуры; происхождение клеточных форм жизни; развитие, изменчивость и видообразование у микроорганизмов и др. До настоящего времени все еще существуют диаметрально противоположные точки зрения на природу вирусов, в том числе и фагов. По мнению одних ученых, фаги относятся к живым организмам; другие рассматривают их как особые вещества типа ферментов. Ва
|
||||
|
|
Последнее изменение этой страницы: 2016-07-16; просмотров: 1239; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.41 (0.021 с.) |