

Мы поможем в написании ваших работ!
ЗНАЕТЕ ЛИ ВЫ?
|
Особенности противовирусного иммунитета
Противовирусный иммунитет начинается со стадии презентации вирусного антигена Т-хелперами.
Иммунитет направлен на нейтрализацию и удаление из организма вируса, его антигенов и зараженных вирусом клеток. Выделяют две основные формы участия антител в развитии противовирусного иммунитета:
1) нейтрализацию вируса антителами;
2) иммунный лизис инфицированных вирусом клеток с участием антител.
| Структурная и молекулярная организация вирусов
| 
| |
|
|
| В онтогенетическом цикле вируса выделены две стадии — внеклеточная и внутриклеточная и, соответственно, две формы его существования — вирион и вегетативная форма. Вирион — это целая вирусная частица, в основном состоящая из белка и нуклеиновой кислоты, часто устойчивая к воздействию факторов внешней среды и приспособленная для переноса генетической информации из клетки в клетку. Вегетативная форма вируса существует в едином комплексе вирус-клетка и только в их тесном взаимодействии.
|
|
| Архитектура вирионов
|
| |
|
|
| Внеклеточная форма вируса — вирион, предназначенная для сохранения и переноса нуклеиновой кислоты вируса, характеризуется собственной архитектурой, биохимическими и молекулярно-генетическими особенностями. Под архитектурой вирионов понимают ультратонкую структурную организацию этих надмолекулярных образований, различающихся размерами, формой и сложностью строения. Для описания архитектуры вирусных структур разработана номенклатура терминов: Белковая субъединица — единая, уложенная определенным образом полипептидная цепь. Структурная единица (структурный элемент) — белковый ансамбль более высокого порядка, образованный несколькими химически связанными идентичными или неидентичными субъединицами. Морфологическая единица — группа выступов (кластер) на поверхности капсида, видимая в электронном микроскопе. Часто наблюдаются кластеры, состоящие из пяти (пентамер) и шести (гексамер) выступов. Это явление получило название пентамерно-гексамерной кластеризации. Если морфологическая единица соответствует химически значимому образованию (сохраняет свою организацию в условиях мягкой дезинтеграции), то применяют термин капсомер. Капсид — внешний белковый чехол или футляр, образующий замкнутую сферу вокруг геномной нуклеиновой кислоты. Кор (core) — внутренняя белковая оболочка, непосредственно примыкающая к нуклеиновой кислоте. Нуклеокапсид — комплекс белка с нуклеиновой кислотой, представляющий собой упакованную форму генома. Суперкапсид или пеплос — оболочка вириона, образованная липидной мембраной клеточного происхождения и вирусными белками. Матрикс — белковый компонент, локализованный между суперкапсидом и капсидом. Пепломеры и шипы — поверхностные выступы суперкапсида. Как уже отмечалось, вирусы могут проходить через самые микроскопические поры, задерживающие бактерии, за что и были названы фильтрующимися агентами. Свойство фильтруемости вирусов обусловлено размерами, исчисляемыми нанометрами (нм), что на несколько порядков меньше, чем размеры самых мелких микроорганизмов. Размеры вирусных частиц, в свою очередь, колеблются в относительно широких пределах. Самые мелкие просто устроенные вирусы имеют диаметр чуть больше 20 нм (парвовирусы, пикорнавирусы, фаг Qβ), вирусы средних размеров — 100-150 нм (аденовирусы, коронавирусы). Наиболее крупными признаны вирусные частицы осповакцины, размеры которых достигают 170x450 нм. Длина нитевидных вирусов растений может составлять 2000 нм. Представители царства Vira характеризуются разнообразием форм. По своей структуре вирусные частицы могут быть простыми образованиями, а могут представлять собой достаточно сложные ансамбли, включающие несколько структурных элементов. Условная модель гипотетического вириона, включающего все возможные структурные образования, представлена на рисунке. Существует два типа вирусных частиц (ВЧ), принципиально отличающихся друг от друга: 1) ВЧ, лишенные оболочки (безоболочечные или непокрытые вирионы); 2) ВЧ, имеющие оболочку (оболочечные или покрытые вирионы).
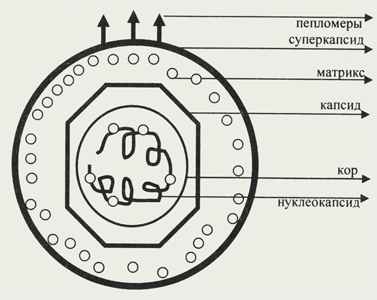 Рис. Строение гипотетического вириона Рис. Строение гипотетического вириона
|
|
|
|
|
|
| | Химический состав вирусов
|
|
| |
|
| | | | | |
|
| Основными химическими соединениями, которые входят в состав всех вирусов, являются белки и нуклеиновые кислоты. В состав ряда вирусов входят липиды и углеводы.
|
| | Белки
|
| |
|
|
| Локализация вирусных белков. Белки, связанные с жизненным циклом вируса, разделяют на белки, детерминируемые геномом вируса и белки, имеющие клеточное происхождение. В качестве примера клеточных белков, которые обнаружены в составе некоторых вирионов, могут быть приведены белок цитоскелета — актин, и ядерные белки — гистоны. Белки клеточного происхождения, участвующие в процессе репликации вируса, будут рассмотрены в разделе взаимодействия вируса с клеткой. По месту локализации белки, детерминируемые вирусным геномом, разделяют на две группы: 1) структурные белки — это белки, входящие в состав ВЧ, их обозначают как VP; 2) неструктурные белки — это предшественники структурных белков, регуляторные белки и ферменты, обслуживающие процесс внутриклеточной репродукции вируса и не входящие в состав ВЧ. Их обозначают как NS-белки (схема). Свойства вирусных белков. В состав вирионов входят белки с различной молекулярной массой (от 4 до 100 кД), состоящие из одной или нескольких полипептидных цепей. Количество этих белков также различно у разных вирусов. В состав нуклеокапсида ВТМ входит один белок. У других вирусов в состав вириона может входить несколько десятков белков, имеющих различные физико-химические свойства. Белки, формирующие капсид, нуклеокапсид и коровую оболочку, обладают одним общим свойством — способностью к самосборке. В состав ВЧ могут входить низкомолекулярные белки, не участвующие в формировании капсида. Например, геномные белки пикорнавирусов и аденовирусов. Геномный белок ковалентно связан с нуклеиновой кислотой и участвует в ее репликации.
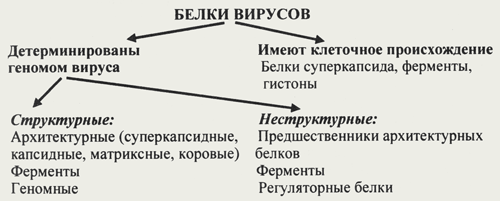 Локализация вирусных белков
Сложные белки представлены гликопротеинами (обозначают как gp) и липопротеинами. Наличие гликопротеина определяет присутствие в вирионе углеводного компонента, который может быть представлен олигосахаридами маннозного типа, галактозой, N-ацетилглюкозамином или нейраминовой кислотой. Вирусные гликопротеины, как правило, экспонированы на наружной поверхности ВЧ и выполняют три основные функции: обеспечивают связывание вириона с клеточным рецептором (функция прикрепительного белка), обладают фузионной активностью (обеспечивают слияние мембран) и определяют антигенные свойства вирусов. В то же время, вирусные гликопротеины могут быть и неструктурными белками и, оставаясь в интегральной форме в мембране шероховатого эндоплазматического ретикулюма (ШЭР), выполнять функции транслоказ, обеспечивая транспорт вирусных компонентов в его просвет. Вирусные липопротеины представлены белками, ацилированными, как правило, миристиновой (C Локализация вирусных белков
Сложные белки представлены гликопротеинами (обозначают как gp) и липопротеинами. Наличие гликопротеина определяет присутствие в вирионе углеводного компонента, который может быть представлен олигосахаридами маннозного типа, галактозой, N-ацетилглюкозамином или нейраминовой кислотой. Вирусные гликопротеины, как правило, экспонированы на наружной поверхности ВЧ и выполняют три основные функции: обеспечивают связывание вириона с клеточным рецептором (функция прикрепительного белка), обладают фузионной активностью (обеспечивают слияние мембран) и определяют антигенные свойства вирусов. В то же время, вирусные гликопротеины могут быть и неструктурными белками и, оставаясь в интегральной форме в мембране шероховатого эндоплазматического ретикулюма (ШЭР), выполнять функции транслоказ, обеспечивая транспорт вирусных компонентов в его просвет. Вирусные липопротеины представлены белками, ацилированными, как правило, миристиновой (C   ) кислотой. Остатки жирных кислот, соединенные с молекулой белка, выполняют функцию липофильного якоря. Вирусные белки-ферменты могут входить в состав вирусной частицы или являться неструктурными белками и появляться в клетке после экспрессии вирусного генома. Наиболее оснащенным ферментами является вирион вируса оспы, который имеет практически полный набор энзимов, необходимых для независимой внутриклеточной репликации вируса. В то же время, мелкие просто организованные изометрические вирусы с позитивным РНК-геномом могут не иметь никаких ферментов в составе вириона. Функционально активные белки вирусов представлены, в первую очередь, ферментами нуклеинового обмена, обеспечивающими сложные механизмы репликации/транскрипции вирусного генома; ферментами, осуществляющими посттрансляционный процессинг и модификацию белков, и ферментами, участвующими в проникновении вирионов в клетку хозяина. Первая группа ферментов наиболее многочисленна и включает как аналоги клеточных ферментов, так и вирус-специфические ферменты. ДНК-зависимая ДНК-полимераза — осуществляет синтез ДНК на матрице ДНК (вирус оспы). ДНК-зависимая РНК-полимераза — осуществляет синтез мРНК на матрице ДНК (вирус оспы). РНК-зависимая РНК-полимераза — осуществляет синтез РНК на матрице РНК. Выполняет функции транскриптазы и репликазы. Впервые обнаружена в 1970 г. Балтимором у вируса везикулярного стоматита. Входит в состав вирионов или является NS-белком РНК-содержащих вирусов. Обратная транскриптаза или ревертаза или РНК-зависимая ДНК-полимераза осуществляет синтез ДНК на матрице РНК. Впервые открыта в 1970 г. у ретровирусов Темином и Мизутани. Хеликаза — осуществляет расплетете двухнитевой структуры ДНК. Кроме этого хеликазы обладают нуклеотидтрифосфат-зависимой РНК-хеликазной активностью, которая включает три процесса: связывание дезоксинуклеотидтрифосфата, его гидролиз и за счет этой энергии расплетение двухнитевой РНК. мРНК-модифицирующие ферменты: поли-А-полимераза — аденилирует 3'-конец РНК за счет энергии АТФ; Кэп-энзим и метилтрансферазный комплекс — катализирует образование на 5'-конце кэп-структуры. АТФ-аза, ГТФ-аза — осуществляют гидролиз соответствующих энергетических субстратов. Рибонуклеаза Н — разрушает РНК, находящуюся в дуплексе с ДНК. Вторая группа вирусных ферментов — ферменты белкового обмена. Здесь мы приведем лишь некоторые из них: Протеиназы — ферменты, участвующие в посттрансляционном процессинге полипротеинов. Являются NS-белками РНК-содержащих вирусов; Протеинкиназы — ферменты, фосфорилирующие структурные белки вирионов. Обнаружены в составе вируса везикулярного стоматита, вируса бешенства, альфавирусов и ретровирусов. Примерами ферментов, участвующих в проникновении вирусов в клетку, являются лизоцим бактериофагов и нейраминидаза вируса гриппа. ) кислотой. Остатки жирных кислот, соединенные с молекулой белка, выполняют функцию липофильного якоря. Вирусные белки-ферменты могут входить в состав вирусной частицы или являться неструктурными белками и появляться в клетке после экспрессии вирусного генома. Наиболее оснащенным ферментами является вирион вируса оспы, который имеет практически полный набор энзимов, необходимых для независимой внутриклеточной репликации вируса. В то же время, мелкие просто организованные изометрические вирусы с позитивным РНК-геномом могут не иметь никаких ферментов в составе вириона. Функционально активные белки вирусов представлены, в первую очередь, ферментами нуклеинового обмена, обеспечивающими сложные механизмы репликации/транскрипции вирусного генома; ферментами, осуществляющими посттрансляционный процессинг и модификацию белков, и ферментами, участвующими в проникновении вирионов в клетку хозяина. Первая группа ферментов наиболее многочисленна и включает как аналоги клеточных ферментов, так и вирус-специфические ферменты. ДНК-зависимая ДНК-полимераза — осуществляет синтез ДНК на матрице ДНК (вирус оспы). ДНК-зависимая РНК-полимераза — осуществляет синтез мРНК на матрице ДНК (вирус оспы). РНК-зависимая РНК-полимераза — осуществляет синтез РНК на матрице РНК. Выполняет функции транскриптазы и репликазы. Впервые обнаружена в 1970 г. Балтимором у вируса везикулярного стоматита. Входит в состав вирионов или является NS-белком РНК-содержащих вирусов. Обратная транскриптаза или ревертаза или РНК-зависимая ДНК-полимераза осуществляет синтез ДНК на матрице РНК. Впервые открыта в 1970 г. у ретровирусов Темином и Мизутани. Хеликаза — осуществляет расплетете двухнитевой структуры ДНК. Кроме этого хеликазы обладают нуклеотидтрифосфат-зависимой РНК-хеликазной активностью, которая включает три процесса: связывание дезоксинуклеотидтрифосфата, его гидролиз и за счет этой энергии расплетение двухнитевой РНК. мРНК-модифицирующие ферменты: поли-А-полимераза — аденилирует 3'-конец РНК за счет энергии АТФ; Кэп-энзим и метилтрансферазный комплекс — катализирует образование на 5'-конце кэп-структуры. АТФ-аза, ГТФ-аза — осуществляют гидролиз соответствующих энергетических субстратов. Рибонуклеаза Н — разрушает РНК, находящуюся в дуплексе с ДНК. Вторая группа вирусных ферментов — ферменты белкового обмена. Здесь мы приведем лишь некоторые из них: Протеиназы — ферменты, участвующие в посттрансляционном процессинге полипротеинов. Являются NS-белками РНК-содержащих вирусов; Протеинкиназы — ферменты, фосфорилирующие структурные белки вирионов. Обнаружены в составе вируса везикулярного стоматита, вируса бешенства, альфавирусов и ретровирусов. Примерами ферментов, участвующих в проникновении вирусов в клетку, являются лизоцим бактериофагов и нейраминидаза вируса гриппа.
|
|
|
|
|
|
| | Липиды
|
|
| |
|
| | | | | |
|
| Клетки всех живых организмов содержат два вида нуклеиновой кислоты — ДНК (двухнитевая ДНК клеточного генома) и РНК (мРНК, тРНК, рРНК). В отличие от клеток, вирионы содержат только один вид нуклеиновой кислоты — ДНК или РНК. И та и другая являются хранителями наследственной информации и выполняют функции генома. Однако следует учитывать, что наличие одного вида нуклеиновой кислоты является характеристикой вириона, но не вируса. В жизненном цикле вируса его геномная нуклеиновая кислота транскрибируется, то есть ДНК-содержащие вирусы образуют РНК. Ряд РНК-содержащих вирусов имеют в цикле репродукции стадию обратной транскрипции и синтезируют ДНК на матрице РНК. Примерно 20% всех вирусов имеют ДНК-геном, 80% — РНК-геном. Способность РНК хранить наследственную информацию — уникальное свойство вирусов. Размеры вирусных геномов (длина нуклеотидных последовательностей, выраженная в нуклеотидах) варьируют в широких пределах — от 1,7 тысяч нуклеотидов (т.н.) у цирковируса свиней до 300 т.н. у фикоднавирусов архибактерий. Кроме того, что геном вирусов может быть представлен или ДНК или РНК, он может находиться в разных видах — в виде двухнитевой (дн) или однонитевой (он) формы, в виде линейной или кольцевой, в виде непрерывной или сегментированной формы (схема).
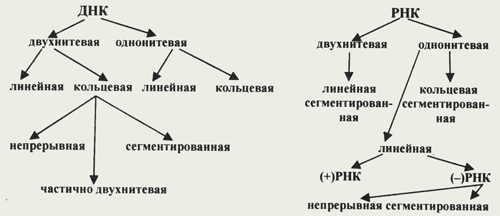 Схема. Виды нуклеиновых кислот вирусов
Особенностями ДНК-геномов является то, что линейные молекулы никогда не имеют бессмысленных концов. Концы молекул могут содержать прямые (>>) или инвертированные концевые повторы (><), выступающие комплементарные (липкие) концы, самокомплементарные концевые последовательности, терминальные геномные белки (O) (рис. 1). Схема. Виды нуклеиновых кислот вирусов
Особенностями ДНК-геномов является то, что линейные молекулы никогда не имеют бессмысленных концов. Концы молекул могут содержать прямые (>>) или инвертированные концевые повторы (><), выступающие комплементарные (липкие) концы, самокомплементарные концевые последовательности, терминальные геномные белки (O) (рис. 1).

| дн, линейная с прямыми концевыми повторами
| Герпесвирусы Фаги T7, T2
| 
| дн, линейная с инвертированными повторами и терминальными белками
| Аденовирусы
| 
| дн, линейная с липкими концами
| Фаг λ
| 
| дн, с ковалентно замкнутыми концами (терминальные петли)
| Поксвирусы Иридовирусы
| 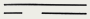
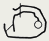
| дн, с разрывами одной цепи
| Фаг T5
| 
| частично дн, кольцевая
| Гепаднавирусы
| 
| дн, кольцевая
| Папиломавирусы Фаг RM2
| 
| дн, кольцевая, сегментированная
| Полиднавирусы
| 
| он, кольцевая
| Фаги φХ174, M13
| 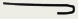
| он, линейная с самокомплементарной 3'-последовательностью
| Парвовирусы
|
Рис. 1. Виды ДНК-геномов вирусов
Многообразие видов РНК геномов расширяется за счет существования последовательностей, отличающихся направлением связей сахаро-фосфатного остова (рис. 2).
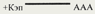
| 5'-кэп, 3'-поли-А последовательность
| Флавирусы Коронавирусы
| 
| 5'-кэп, 3'-тРНК-подобная структура
| ВТМ
| 
| 5'-терминальный геномный белок
| Калицивирусы Пикорнавирусы Потивирусы
| 
| диплоидный набор
| Ретровирусы
| 
| он, линейная
| Парамиксовирусы Рабдовирусы
| 
| он, линейная, сегментированная
| Ортомиксовирусы (7-8 сегментов)
| 
| он, кольцевая, сегментированная
| Буньявирусы Аренавирусы
| 
| дн, линейная, сегментированная
| Реовирусы (10-11 сегментов)
| 
| обоюдозначащая (амбисенс) РНК
| S-сегмент аренавирусов Тосповирусы
|
Рис. 2. Виды РНК-геномов вирусов
Однонитевые РНК могут иметь позитивную полярность — (+)РНК, негативную полярность — (-)РНК или могут быть представлены обоюдозначащей цепью — (+,-)РНК (амбисенс стратегия кодирования). В свою очередь, РНК позитивной полярности могут иметь разную структурную организацию: могут, являясь матричной РНК, иметь на 5'-конце кэп (7-метилгуанозин, Сар), а на 3'-конце — поли-А (poly-A) последовательность; могут не иметь кэпа или поли-А; могут иметь на 5'-конце геномный белок; могут иметь на 3'-конце тРНК-подобную или шпильковую структуру.
Виды геномов вирусов легли в основу их классификации. Однако следует учитывать, что вид генома в настоящее время не является формальным таксоном и используется для удобства ориентации в многообразии вирусов.
|
| | Проникновение вирусов в организм хозяина
|
| |
|
|
| Циркулируя в биосфере, вирус встречает восприимчивого хозяина, проникает в организм и может вызвать инфекцию. Инфекция (infectio) — заражение. Как биологическое явление представляет собой процесс взаимодействия одного или нескольких видов возбудителей-паразитов с более высокоорганизованным организмом хозяина, который может проявляться болезнью или носительством. Совокупность вызываемых патологических изменений в организме определяется как инфекционный процесс. Проникновение вируса в организм хозяина у разных биологических видов решается по-разному. 1. Вирусы растений проникают в организм хозяина по типу раневых инфекций, где распространяются по плазмодесмам, ксилеме и флоэме. 2. Вирусы бактерий — путем введения нуклеиновой кислоты в тело клетки или путем проникновения вириона. 3. Вирусы насекомых попадают в организм хозяина в процессе питания или размножения. 4. Вирусы животных и человека при инфицировании организма хозяина проходят более сложный путь. Одни вирусы (вирус гриппа, ротавирусы) реплицируются и вызывают заболевание в месте проникновения в организм (входные ворота инфекции). Другие вирусы, попав в организм хозяина с использованием того или иного механизма, проходят стадию распространения. Распространение вируса в организме сопровождается виремией (вирусемией) — циркуляцией вируса в крови, что свидетельствует о генерализации инфекции. Различают несколько путей распространения вирусов в организме: 1. Нейронный путь (вирусы бешенства, герпеса). 2. Лимфатический путь (реовирусы, полиомавирусы). 3. Гематогенный путь, ассоциированный с клеточными компонентами и плазмой крови (вирус краснухи, вирусы гепатита B и C, цитомегаловирус, энтеровирусы). Сохранение вируса как биологического вида обеспечивает его восприимчивый хозяин, который является основным элементом экологической ниши вируса. Способность клеток или организма хозяина заражаться называется восприимчивостью. Спектр хозяев разных вирусов значительно варьирует. Одни вирусы имеют широкий круг хозяев, другие заражают лишь определенные клетки одного вида хозяина. Широта круга хозяев может быть ограничена видом (видоспецифические вирусы) или определяться таксономическими категориями более высокого порядка. Вирусы, имеющие широкий круг хозяев, распространены среди вирусов растений. Например, вирус табачной мозаики и вирус желтухи астр инфицируют как растения, так и своего переносчика-насекомого. Примером вирусов животных и человека, имеющих несколько хозяев, являются арбовирусы. Вирус лихорадки Западного Нила инфицирует человека, комаров, водоплавающих птиц; вирус клещевого энцефалита — человека, животных, клещей. Однако не найдены вирусы, способные поражать одновременно клетки прокариот и эукариот. Экологические ниши вирусов, выражающиеся в круге восприимчивых хозяев, сложились в ходе эволюции. Однако они могут меняться, и вирусы способны преодолевать межвидовые барьеры. Так, вирусы гриппа циркулируют среди многих видов животных и птиц, которые являются резервуаром сохранения вируса в межэпидемический период. В Китае среди детей часто наблюдаются вспышки кишечной инфекции, вызванной ротавирусами антигенной группы B, резервуаром которых являются крысы. В качестве примера вируса, преодолевшего межвидовой барьер, можно привести вирус иммунодефицита человека (ВИЧ), который перешел в человеческую популяцию от обезьян. Процесс адаптации вируса к новым хозяевам определяется мутациями рецептор-распознающих участков вирионов и собственно изменением вирулентности для данного биологического вида. Однако следует понимать, что не все так однозначно и изучение молекулярных основ преодоления межвидовых барьеров еще впереди. Для того чтобы размножиться, вирус должен найти восприимчивую клетку. Каждый вирус обладает так называемой тканевой тропностью — способностью инфицировать клетки определенного типа. Так, вирусы растений поражают или ткань листа, или ткань прицветника или клетки корневой системы. Вирусы бактерий видоспецифичны — вирусы архибактерий не могут инфицировать клетку E. coli, а многие колифаги не проникают в клетку шигеллы. Наиболее выражена тканевая специфичность вирусов животных и человека. Так, вирусы гепатитов поражают гепатоциты, вирус Эпштейна-Барр (вызывает инфекционный мононуклеоз) обладает тропностью к B-лимфоцитам, ВИЧ — к T-лимфоцитам, кишечные вирусы — к энтероцитам, кардиотропностью обладают вирусы Коксаки B. Целый ряд вирусов обладает тропностью не к одному, а к нескольким типам клеток. Так полиовирусы тропны к клеткам респираторного тракта, желудочно-кишечного тракта (ЖКТ), центральной нервной системы (ЦНС). Вирус гепатита C (ВГC) лимфотропен и гепатотропен. Специфическое сродство вирусов к клеткам и тканям определяется двумя механизмами: — Присутствием на клеточной поверхности специфических для вируса рецепторов. — Содержанием в системе активирующих ферментов, необходимых для протеолитического расщепления вирусных поверхностных белков и проявления инфекционной активности вируса.
|
| | Взаимодействие вирусов с клеткой
|
| |
|
|
| Основные этапы взаимодействия: адсорбция вируса на клеточной поверхности; проникновение-раздевание вируса; реализация генетической информации; морфогенез вирусных частиц; выход вирусов из клетки.
|
|
|




