Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Организация и функционированиеСодержание книги
Поиск на нашем сайте
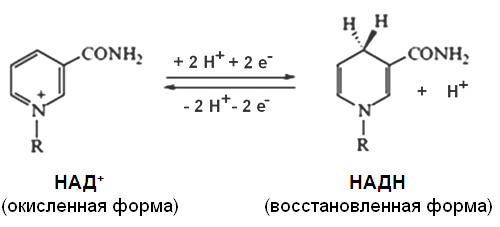
ДЫХАТЕЛЬНОЙ ЦЕПИ Окисление субстратов в процессе дыхания можно представить как перенос электронов и протонов (т. е. атомов водорода) от органических веществ на кислород. В этом процессе участвует ряд промежуточных переносчиков, образующих дыхательную цепь. Дыхательная цепь (электронотранспортная цепь, цепь переноса электронов) - система трансмембранных белков и переносчиков электронов, которые передают электроны от субстратов на кислород. В клетках эукариот дыхательная цепь расположена во внутренней мембране митохондрий. При взаимодействии НАД+ и НАДФ+ с атомами водорода происходит обратимое присоединение атомов водорода. В молекулу НАД+ (НАДФ+) включаются 2 электрона и один протон, второй протон остается в среде:
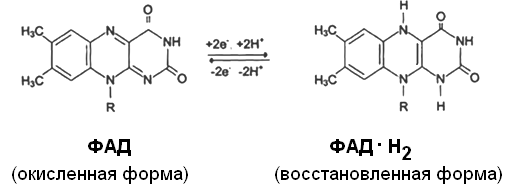
Другим первичным источником атомов водорода и электронов служит восстановленный флавопротеин (ФАД или ФМН):
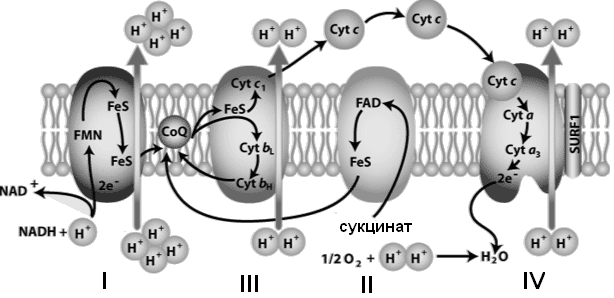
Восстановленные формы этих кофакторов способны транспортировать водород и электроны к дыхательной цепи митохондрий. Компоненты дыхательной цепи встроены в митохондриальную мембрану в виде 4 белково-липидных комплексов (рис. 33).
Комплекс I (НАДН-дегидрогеназа) включает ФМН и железосерный белок FeS (негемовое железо). Железосерный белок участвует в окислительно-восстановительном процессе. Комплекс I окисляет НАДН, перенося с него 2 электрона на кофермент Q (KоQ) и перекачивает 4 протона из матрикса в межмембранное пространство митохондрии. KoQ (убихинон) - производное бензохинона. Это некрупная липофильная молекула. Перемещаясь в липидном слое мембраны, убихинон обеспечивает передачу электронов между комплексами I - III и II - III. Комплекс II (сукцинат-дегидрогеназа) включает ФАД и железосерный белок. Обеспечивает вход в цепь дополнительных электронов за счет окисления сукцината. Комплекс III (QН2-дегидрогеназа) включает цитохромы b и с1 и железосерный белок. Цитохромы - гемопротеины, в которых простетическая геминовая группа близка к гему гемоглобина (у цитохрома b идентична). Комплекс III переносит электроны с убихинона на цитохром с и перекачивает Комплекс IV (цитохром c оксидаза) состоит из цитохромов a и a3, которые, помимо гема, содержат ионы меди. Комплекс IV катализирует перенос электронов с молекул цитохрома на O2 и перекачивает 4 протона в межмембранное пространство.
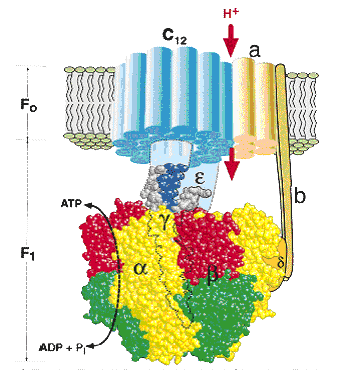
Цитохром а3 - терминальный участок дыхательной цепи (цитохромоксидаза): происходит окисление цитохрома с и образование воды. В организме человека митохондриальная дыхательная цепь образует 300-400 мл воды за сутки (метаболическая вода). Компоненты дыхательной цепи митохондрий расположены в порядке убывания окислительно-восстановительного потенциала. Перемещение электронов в дыхательной цепи происходит по градиенту окислительно-восстановительного потенциала и является источником энергии для переноса протонов. Перенос двух электронов через каждый комплекс обеспечивает перекачку четырех протонов. В результате по сторонам мембраны возникает разность концентраций протонов и одновременно разность электрических потенциалов со знаком «плюс» на наружной поверхности. Электрохимический потенциал понуждает протоны двигаться в обратном направлении - с наружной поверхности внутрь. Однако мембрана непроницаема для них, за исключением участков, где располагается фермент протонная АТФ-синтаза (рис. 34). АТФ-синтаза состоит из двух частей – статора и ротора. Статор состоит из трех α-субъединиц и трех β-субъединиц – они участвуют непосредственно в синтезе АТФ из АДФ и фосфата. К ним примыкает δ-субъединица, и все вместе они образуют F1-субъединицу. Ротор состоит из g- и e-субъединиц. Статор держится в мембране, а ротор вращается за счет энергии протонов. В статоре имеется протонный канал (F0). Он состоит из двух полуканалов, которые смещены один относительно другого. Протон проходит одну половину канала, затем на вращающемся роторе попадает во вторую половину канала.
Движущей силой для АТФ-синтазы, катализирующей реакцию АДФ + Н3РО4 = АТФ + Н2О, является разность электрохимических потенциалов, создаваемая при движении протонов через канал. П. Митчелл для объяснения молекулярного механизма сопряжения транспорта электронов и образования АТФ в дыхательной цепи в 1960 г. предложил хемиосмотическую концепцию:в дыхательной цепи есть только 3 участка (комплексы I, III, IV), где перенос электронов сопряжен с накоплением энергии, достаточным для образования АТФ.
Коэффициент фосфорилирования - отношение величины образовавшейся АТФ к поглощенному кислороду: АТФ/О или Р/О. Максимальная величина коэффициента фосфорилирования 3, если реакция окисления идет с участием НАДН+Н+, и 2, если окисление субстрата протекает через ФАДН2. Реально получаемые величины меньше (2,5 и 1,5), т.е. процесс дыхания не полностью сопряжен с фосфорилированием. Степень сопряжения зависит главным образом от целостности митохондриальной мембраны. Образующаяся АТФ при участии АДФ-АТФ-транслоказы транспортируется из матрикса на наружную сторону мембраны и попадает в цитозоль. Одновременно та же транслоказа переносит АДФ в обратном направлении, из цитозоля в матрикс митохондрии. Общее содержание АТФ в организме 30-50 г, но средняя продолжительность жизни молекулы АТФ меньше 1 мин. В сутки у человека синтезируется 40-60 кг АТФ и столько же распадается. На каждое сокращение сердечной мышцы расходуется около 2% имеющейся в ней АТФ. Вся АТФ израсходовалась бы за 1 мин., если бы не было ее регенерации. При образовании тромба в коронарной артерии поступление кислорода в клетки прекращается, соответственно прекращается и регенерация АТФ, и клетки погибают (инфаркт миокарда). Увеличение концентрации АДФ приводит к ускорению дыхания и фосфорилирования. Зависимость интенсивности дыхания митохондрий от концентрации АДФ называют дыхательным контролем. Для оценки влияния адениловых нуклеотидов на процессы метаболизма используют энергетический заряд клетки (ЭЗК):
В норме ЭЗК = 0,7-0,8: скорость образования АТФ равна скорости ее использования, адениловая система насыщена энергией. При ЭЗК < 0,7 ускоряется образование АТФ путем увеличения скорости реакций общего пути катаболизма. Если ЭЗК = 1, то процессы синтеза АТФ тормозятся и ускоряется ее использование. Механизм дыхательного контроля характеризуется высокой точностью. Относительные концентрации АТФ и АДФ в тканях изменяются в узких пределах, тогда как потребление энергии клеткой может изменяться в десятки раз. Т.о., энергия пищевых веществ в клетке трансформируется сначала в энергию АТФ, а затем АТФ служит непосредственным источником энергии для биохимических и физиологических процессов. Эти превращения энергии и есть энергетический обмен. Гипоэнергетические состояния подразделяются на: 1. Алиментарные (голодание, авитаминоз). 2. Гипоксические. Связаны: - с нарушением поступления кислорода в кровь. Экзогенная гипоксия -недостаток кислорода во вдыхаемом воздухе, легочная (дыхательная) –нарушение легочной вентиляции; - с нарушением транспорта кислорода в крови. Гемодинамическая гипоксия связана с нарушениями кровообращения (генерализованные – пороки сердца, кровопотеря; локальные – спазм сосудов, тромбоз); причины гемоглобиновой гипоксии – гипогемоглобинемия, гемоглобинопатии, блокирование гемоглобина ядами. 3. Митохондриальные. Затруднено использование кислорода в клетках в результате нарушения функций митохондрий ингибиторами ферментов дыхательной цепи, разобщителями окисления и фосфорилирования, мембранотропными веществами. При полном голодании пищевых резервов организма хватает на несколько недель. При лишении же организма кислорода смерть наступает через 2-3 минуты. Поэтому гипоксия - наиболее частая причина гипоэнергетических состояний, а гипоксия мозга - непосредственная причина смерти. Среди реанимационных процедур ведущее место занимают меры, направленные на восстановление снабжения органов кислородом.
|
|||||||||||
|
|
Последнее изменение этой страницы: 2016-07-14; просмотров: 226; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.147.27.154 (0.012 с.) |