Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Различие понятий «законы раздражения» возбудимых тканей и «законы возбуждения»Содержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте Различие понятий «законы раздражения» возбудимых тканей и «законы возбуждения»
Не следует путать «законы раздражения» возбудимых тканей и «законы возбуждения»
Законы раздражения отвечают на вопрос, каким должен быть раздражитель, чтобы возникло возбуждение. Законы возбуждения отвечают на вопрос, каким образом может ответить возбудимая структура на действие раздражителя.
К законам раздражения относятся законы: 1. силы 2. времени 3. градиента силы
К законам возбуждения относятся законы: 1. все или ничего 2. силы
Не путайте закон «силы», как закон возбуждения, с законом «силы», как законом раздражения. Сравните: Определение «закона силы» (возбуждения): с увеличением силы стимула увеличивается сила ответной реакции возбудимой структуры. Определение «закон «силы» (раздражения): чтобы возникло возбуждение, стимул должен быть достаточно сильным – пороговым или выше порогового[Б4]. Область применения «закона силы» (возбуждения) - описание процесса возбуждения. Область применения «закон «силы» (раздражения) - характеристика стимула. Так при выполнении закона «силы» раздражения -– возникает возбуждение, которое в свою очередь может протекать или по закону «силы» или по закону «всё или ничего».
Понятия «раздражитель», «раздражение» в физиологии возбудимых тканей
Раздражитель [Б5] - фактор внешней или внутренней по отношению к возбудимой структуре среды, который при действии или изменении действия, способен вызвать возбуждение.
Естественно, речь идёт об определении понятия раздражитель в контексте физиологии возбудимых тканей. Напомню, на действие раздражителя (стимула) структура может ответить раздражение (неспецифической реакцией) и возбуждением (специфической электрической реакцией [V.G.6]). Возбуждение возникает при выполнении соответствующих законов раздражения. Для реакции раздражения в тех же возбудимых структурах выполнение рассматриваемых сегодня нами законов совершенно не обязательно. Ответить на раздражение возбуждением могут только возбудимые ткани, их составляющие и органы из них состоящие. Например, мышечное волокно, мышечная ткань, мышца (орган). Напомню, к возбудимым тканям относят нервную, мышечную и железистую.
Всё чаще вместо термина «раздражитель» применяется термин «стимул». Это синонимы. И мы в дальнейшем термин стимул будем применять очень часто. Но запомните! В физиологии возбудимых тканей есть понятие возбуждение, но нет понятия возбудитель. Возбуждение возникает на действие раздражителя (стимула).
Итак, согласно определению раздражителем может быть фактор, который ранее не действовал на возбудимую структуру. Например, Вашей руки коснулся сосед. Если Вы это почувствовали, в определённых возбудимых структурах возникло возбуждение.
Другой пример. В рецепторах, контролирующих газовый состав крови, возбуждение возникает при изменении концентрации кислорода или углекислого газа в крови.
Может ли возникнуть возбуждение без внешнего стимула? Да, в результате спонтанной деполяризации клетки. Эти процессы характерны для клеток‑пейсмекеров сердечной мышцы и желудочно-кишечного тракта.
Типы раздражителей
Признаки, по которым различаются раздражители: 1. Природе (модальность, валентность): физические, химические и т.п. 2. Биологическому значению (адекватные, неадекватные) 3. Отношению силы воздействия к порогу [V.G.7] возбуждения (подпороговые, пороговые, сверхпороговые). 4. Одиночные или серийные
По природе [Б8] раздражители разделяют на химические, механические, лучистые, температурные, электрические и т.д.[Б9]. В этом случае говорят о модальности[Б10] стимула[A11]. Стимулы одной и той же модальности различаются по валентности. Например, химические (модальность) раздражители могут быть солёными, сладкими, горькими, кислыми (валентность).ермин модальность, чаще применяют в области сенсорной физиологии касательно рецепторов и анализаторов в целом[Б12]. И когда говорят о модальности раздражителя, имеют в виду характер вызываемых раздражителем ощущений. Но не забудем, что рецепторы, да и анализаторы в целом – это возбудимые структуры. Внутри каждой модальности можно выделить валентность раздражителя. Например, химический раздражитель может быть кислотой, щелочью, солью[Б13].
По биологическому значению независимо от модальности раздражители делят на адекватные и неадекватные[Б14]. Адекватные раздражители способны при воздействии на определенные возбудимые [V.G.15] структуры вызвать реакцию возбуждения. Другими словами, раздражитель, действуя на разные биологические структуры, может вызвать возбуждение только в некоторых из них. Вот для этих структур этот раздражитель будет адекватен. Например, действие света, только в определённых структурах сетчатки глаза вызывает возбуждение. Для них он адекватен. Необязательно, говоря об адекватных раздражителях, замыкаться в рамках «естественных условий» и отождествлять понятия «естественный раздражитель» и «адекватный раздражитель». Например, действие на вкусовые рецепторы химических веществ пищи вызывает возбуждение. Химические вещества пищи, безусловно, в этом случае являются и естественными и адекватными раздражителями. Но, если мы в лабораторных условиях подействуем на эти же рецепторы электрическим током, может также возникнуть возбуждение. В этом случае раздражитель никак не будет естественным, но будет адекватным для рассматриваемых рецепторов. Процитируем другое определение адекватных раздражителей. «Адекватные раздражители — это такие раздражители, которые воздействуют в естественных условиях на строго определенные рецепторы и возбуждают их[Б16] [++484+ с238]». Вы должны понять, почему приводимое определение, по меньшей мере, неточно.
Неадекватные раздражители способны при воздействии на определенные возбудимые [V.G.17] структуры вызвать реакцию возбуждения, но при этом необходимы затраты энергии существенно большие, чем при возбуждении этих же структур от адекватного раздражителя. Например, видимый свет для рецепторов сетчатки или звук в диапазоне его восприятия для рецепторов слухового анализатора является адекватным раздражителем[Б18]. Однако ощущение вспышки света (фосфен, «искры из глаз») или слышимого звука (звона в ушах) может возникнуть при действии механических (удар по голове) и других раздражителей достаточной силы[Б19]. В данном случае также возникает возбуждение соответственно в зрительном или слуховом анализаторах, но уже под влиянием не свойственных для них неадекватных раздражителей[Б20]. Адекватность раздражителя проявляется в том, что его пороговая сила значительно ниже по сравнению с пороговой силой неадекватного раздражителя[Б21]. Например, ощущение света возникает у человека, когда минимальная интенсивность светового раздражителя составляет всего 10-17 - 10-18 Вт, а механического – более. 10-4 Вт, т.е. разница между световым и механическим пороговым раздражителями для рецепторов глаза человека достигает 13-14 порядков. Ещё раз подчеркну, неадекватные раздражители тоже способны вызвать возбуждение. Когда мы говорим о неадекватных раздражителях для какой‑либо возбудимой структуры, имеем ввиду, что для этой же структуры имеются адекватные раздражители.
Может ли стимулы одной и той же модальности, но разной валентности различаться по адекватности возбудимой структуре? Да, могут. Например, такие химические (модальность) раздражители как сахар, соль (валентность) являются адекватными для разных вкусовых рецепторов языка.
По отношению силы воздействия раздражителя к порогу [V.G.22] возбуждения различают подпороговые, пороговые, сверхпороговые. Подробнее об этой важнейшей характеристике раздражителя мы будем говорить позже, разбирая «закон силы» раздражения.
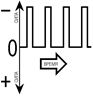
Раздражители могут быть одиночные и серийные. Одиночные раздражители различаться по силе, длительности, форме, скорости нарастания и уменьшения силы (градиенту) (рис. 809141947).
Рис[V.G.23]. 809141947. Различие параметров одиночных раздражителей (стимулов): а — по силе, b — по длительности, c — по скорости нарастания силы (градиенту), d — по форме (первый – прямоугольный, два последующих – трапецевидные[V.G.24]). Серийные раздражители различаться по частоте, меандру (паттерну, рисунку) (рис.).
Рис[Б25].. Различие параметров серийных раздражителей (стимулов): А — по частоте, B — по соотношению продолжительности стимула к продолжительности паузы (скважности), C — по характеру и порядку следования импульсов (меандру).
Обратите внимание, все вышеперечисленные характеристики относятся к раздражителям любой модальности.
Внимание! Таких стимулов, которые нередко изображают студенты, быть не может.
Рис.. Определение скважности. Объяснения в тексте.
Рис.. Однополярный меандр со скважностью 3. Объяснения в тексте.
Рис.. Место расположения и полярность раздражающего электрода. Объяснение в тексте.
Другими словами возбуждение может возникнуть только в месте прохождения токов выходящего направления (рис.).
Рис.. Направление деполяризующих выходящих (А, B) и гиперполяризующих входящих токов (C, D). Объяснение в тексте.
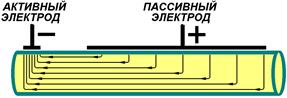
Эффективность раздражения определяется не только абсолютным значением тока, но и плотностью тока под стимулирующим электродом. Плотность тока определяется отношением величины тока, протекающего по цепи, к величине площади электрода. Поэтому при монополярном раздражении площадь активного электрода всегда меньше пассивного (рис.)[Б30].
Рис.. Соотношение площадей активного и пассивного электродов. Объяснение в тексте.
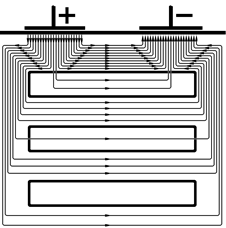
Недостаток внеклеточного подведения тока заключается в значительном ветвлении его тока: только часть его проходит через мембраны клеток, часть же ответвляется в межклеточные щели. Вследствие этого при раздражении приходится применять ток значительно большей силы, чем необходимо для возникновения возбуждения.
Рис. 210041846. Ветвление тока в ткани при раздражении через наружные (внеклеточные) электроды (схема). Волокна возбудимых клеток (ткани) обведены толстой линией, между ними – межклеточные щели [++421+c62]. Объяснение в тексте. По вышеперечисленным причинам и в первом опыте Гальвани лучше наносить стимул биметаллическим пинцетом на нерв
Рис.. Направление деполяризующих выходящих (А, B) и гиперполяризующих входящих токов (C, D). Объяснение в тексте.
При внутриклеточном способе подведения тока к клеткам —– микроэлектрод вводят в клетку, а пассивный электрод прикладывают к поверхности ткани. В этом случае весь ток проходит через мембрану клетки, что позволяет точно определить наименьшую силу тока, необходимую для возникновения потенциала действия. При таком способе раздражения отведение потенциалов производят с помощью второго внутриклеточного микроэлектрода. Пороговая сила тока, необходимая для возникновения возбуждения различных клеток при внутриклеточном раздражающем электроде, равна 10-7 ‑ 10-9 А.
Закон силы
Прежде всего, необходимо помнить, что возбуждение может возникнуть при деполяризации мембраны до критического уровня (КУД). Раздражитель минимальной силы, вызывающий возбуждение называется пороговым. Раздражитель, сила которого превышает пороговый уровень, называется сверхпороговым. Следует обратить внимание на то, что, чем больше сила сверхпорогового раздражителя, тем быстрее возникает возбуждение. Введение понятия «порог раздражения» как следствия закона силы является очень важным для оценки возбудимости объекта[Б33].
Рис. 1306000041. Закон силы. A ‑ подпороговый стимул, B – пороговый стимул, C – сверхпороговый стимул. Вверху – изменения мембранного потенциала при раздражении, внизу – раздражающие стимулы. ПП – уровень мембранного потенциала покоя, КУД – критический уровень деполяризации (порог). ПД – потенциал действия. Объяснения в тексте.
Закон времени
Следует подчеркнуть, что согласно закону времени, слишком короткие по длительности стимулы не способны вызвать возбуждение, какими бы сильными они небыли. Это используется в физиотерапии при получении калорического эффекта при воздействии токами высокой частоты[Б34].
Рис. 209271137. Закон времени A — подпороговый стимул достаточной длительности для возникновения потенциала действия, B — пороговый стимул, не достаточной длительности для возникновения потенциала действия C — подпороговый стимул более чем достаточной длительности для возникновения потенциала действия. Вверху – изменения мембранного потенциала при раздражении, внизу – раздражающие стимулы. ПП – уровень мембранного потенциала покоя, КУД – критический уровень деполяризации (порог). ПД – потенциал действия. Объяснения в тексте.
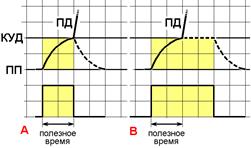
Важным следствием закона времени является введение понятия полезное время – минимальное время, которое необходимо для действия рассматриваемого раздражителя, чтобы возникло возбуждение. Почему полезное? Потому что дальнейшее действие раздражителя на структуру в состоянии возбуждения бесполезно, ничего уже не изменишь. Бесполезно теряется время.
Рис. 209271409. Полезное время при стимулах разной длительности. A ‑ стимул по длительности равен полезному времени, B – стимул по длительности больше полезного времени. Вверху – изменения мембранного потенциала при раздражении, внизу – раздражающие стимулы. ПП – уровень мембранного потенциала покоя, КУД – критический уровень деполяризации (порог). ПД – потенциал действия. Объяснения в тексте.
Чем больше сила раздражителя, тем меньше полезное время. Но об этом чуть позже.
Закон градиента
Рис.. Закон градиента. Рис. 809150447. Кривая «сила – время» Гоорвега-Вейса-Лапика
Сначала для дозирования времени воздействия электрического тока на ткань был применен пистолет, пуля которого могла замыкать и размыкать контакты электрической цепи в течение короткого интервала времени, а затем, для регулирования длительности раздражения, стал использоваться заряд конденсаторов различной емкости[Б41]. Известно, что время разряда конденсатора определяется величиной его емкости и сопротивлением цепи разряда[Б42]. Упомянутые методические приемы позволили Л.Лапику наносить раздражения очень короткой длительности (до 0,001 с и меньше) и исследовать зависимость ответной реакции от силы и длительности раздражения[Б43].
Кривые «силы-времени» хорошо характеризуют возбудимость объектов. Очевидно, возбудимость структуры 1 больше, чем 2.
Рис.. Сравнение возбудимости двух возбудимых структур.
Хронаксия, хронаксиметрия [Б44] Для числовой характеристики экспериментально полученных зависимостей часто используют показатель называемый хронаксией. Хронаксия (от греч. chrónos — «время» и axía — «цена», «мера») — полезное время [Б45] раздражения, сила которого равна удвоенной реобазе минимальное время[A46] [Б47]. Понятие "хронаксия" введено французским физиологом Луисом Лапиком в 1909 году.
Рис[V.G.48]. 209272000. Зависимость между силой тока и временем его действия. Хронаксия (по Гоорвегу, Вейсу и Лапику). Р — реобаза, Хр — хронаксиия.
При хронаксиметрии вначале определяется реобаза, т.е. пороговая сила раздражения при достаточно большой его длительности[Б49]. Время, в течение которого действует или должен действовать пороговый раздражитель, равный значению реобазы, получило название полезного времени[Б50]. Определив реобазу, производится удвоение найденного значения и находится минимальная длительность, при которой это электрическое раздражение способно вызвать возбуждение и ответную реакцию[Б51] - хронаксия. Хронаксия нервных и поперечнополосатых скелетных мышечных волокон человека равна тысячным и десятитысячным долям секунды[Б52]. У гладких мышечных волокон она значительно больше[Б53]. Измерение хронаксии - хронаксиметрия - применяется для изучения работы нервного и двигательного аппарата человека и животных. Проводится с помощью специальных приборов хронаксиметров[Б54]. В клинической практике метод хронаксиметрии применяется в диагностических целях и для изучения закономерностей патологических процессов[Б55]. Внимание! Часто студенты пишут «хрон о ксия». Правильно «хрон а ксия».
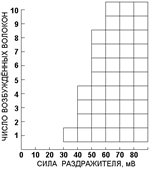
Рис.. Графические способы отображения законов возбуждения. Для одиночных образований (нерное волокно, мышечное волокно) выполняется закон «всё или ничего[Б57]». Если речь идет о целом образовании, например, нервном стволе, содержащем отдельные аксоны, или о скелетной мышце как совокупности отдельных мышечных волокон, то в этом случае каждое отдельное волокно тоже отвечает на раздражитель по типу "все или ничего", но если регистрируется суммарная активность объекта (например, внеклеточно отводимый ПД), то его амплитуда в определенном диапазоне находится в градуальной зависимости [Б58] от силы раздражителя: чем больше сила раздражителя, тем больше ответ. Пример: пусть имеется нервный ствол, состоящий из 10 аксонов. Пороги раздражения для них таковы: 30 мВ - 1-й, 40 мВ - 2, 3, 4-й, 50 мВ - 5, 6, 7, 8-й и 60 мВ - 9 и 10-й аксоны. Следовательно, при 30 мВ - активируется 1 аксон, при 40 мВ - 4 (1-й-+-2, 3, 4-й), при 50 мВ - 8 (1-й+2, 3, 4-й + 5, 6, 7, 8-й), а при 60 мВ - все 10 волокон.
Рис.. Градуальная зависимость между силой раздражения нервного ствола и числом возбужденных нервных волокон. Объяснение в тексте. Рис. 210041815. Закон силы раздражения в приложении к составной возбудимой структуре (нерву, мышце). 6. Действие постоянного подпорогового тока на возбудимые структуры[Б62] [a63]
В 1859 г. немецкий физиолог Пфлюгер Э.Ф.В. установил, что если на нерв воздействовать слабым (подпороговым) постоянным током, то его возбудимость под катодом повышается, а под анодом снижается.
В 1883 г. российский (пермский) физиолог Б.Ф.Вериго значительно дополнил наблюдения Э.Пфлюгера и показал, что как повышение возбудимости под катодом, так и снижение её под анодом характерно только для первоначального действия постоянного подпорогового тока, т.е. это явление временное. Если ток действует достаточно долго[Б65], то под катодом возбудимость снижается, становясь меньше исходной (в состоянии покоя), а под анодом может повыситься.
Как это объясняют? Разберём механизм действия постоянного подпорогового тока на возбудимые структуры в рамках мембранной теории возбуждения. Вначале выясним вопрос как располагаются электроды, через которые на возбудимую структуру подаётся подпороговый ток. Раздражающие электроды могут быть расположены внеклеточно (рис. 209220945) и внутриклеточно (рис. 209220946). [A67]
Рис. 209220945. Схема опыта по влиянию постоянного подпорогового тока на возбудимость при внеклеточной «аппликации тока».
Рис. 209220946. Схема опыта по влиянию постоянного подпорогового тока на возбудимость при внутриклеточной «инъекции тока».
При внеклеточном расположении электродов говорят об «аппликации тока», при внутриклеточном — об «инъекции тока [A68] ». У одного и другого способа воздействия есть достоинства и недостатки[V.G.69]. При «инъекции тока» по сравнению с «аппликацией тока» все будет наоборот: то, что происходит при аппликации под катодом, будет происходить при инъекции анода, а то, что происходит при аппликации под анодом, будет происходить при инъекции катода. Мы подробно рассмотрим действие тока при его аппликации (внеклеточном расположении электродов), как это делали классики Э.Пфлюгер и Б.Ф.Вериго.
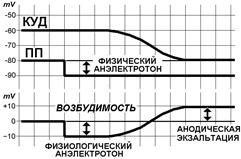
Вначале действия постоянного тока под катодом происходит деполяризация мембраны (физический катэлектротон), а под анодом — гиперполяризация (физический анэлектротон) (рис. 209192100). Для облегчения понимания разбираемых явлений введём конкретные числовые значения величин. На рис. 209192100 под катодом уровень мембранного потенциала поднялся с ‑80 мВ (потенциал покоя) до ‑70 мВ (состояние деполяризации). Под анодом катодом уровень мембранного потенциала снизился с ‑80 мВ (потенциал покоя) до ‑90 мВ (состояние гиперполяризации). Не будем забывать, что если уровень мембранного потенциала изменился от –80 до‑70 мВ говорят о его уменьшении, а с –80 до –90 мВ — о его увеличении. При этом вначале действие постоянного тока уровень критической деполяризации или не изменяется, или его изменения малы по сравнению со сдвигами мембранного потенциала. Следовательно, мембранный потенциал под катодом приближается, а под анодом удаляется от критического уровня деполяризации. Значит под катодом порог раздражения уменьшается на 10 мВ и возбудимость растёт, а под катодом увеличивается на 10 мВ и возбудимость уменьшается. Не забыли, что такое порог раздражения? Это критический уровень деполяризации (критический потенциал) минус мембранный потенциал (КУД-МП). При длительном действии постоянного тока, как и при воздействии медленно нарастающих по силе раздражителей, происходит сдвиг критического уровня деполяризации (КУД). При этом направленность сдвига критического уровня деполяризации и под катодом и под анодом соответствует изменению мембранного потенциала, а абсолютная величина сдвига будет больше. Это в конечном итоге приводит к снижению возбудимости под катодом (катодическая депрессия), а под анодом к возможному её повышению (анодическая экзальтация) (рис. 209192100).
Рис.. Изменение электрофизиологических параметров возбудимых структур при действии постоянного подпорогового тока. КУД – критический уровень деполяризации, ПП – потенциал покоя, МП – мембранный потенциал. Возбудимость выражена в единицах преодоления порога раздражения в состоянии покоя (КУД-ПП). Стрелкой отмечено начало действия тока.
В нашем примере (рис. 209192100) уровень КУД под катодом повышается с –60 мВ до –40 мВ. Значит, порог раздражения становится равным 30 мВ. Т.е. он больше порога раздражения в состоянии покоя клетки на 10 мВ. Под анодом в нашем примере (рис. 209192100) уровень КУД снижается с –60 мВ до –80 мВ. Значит, порог раздражения становится равным 10 мВ. Т.е. он меньше порога раздражения в состоянии покоя клетки на 10 мВ.
Ещё раз повторим введенные выше понятия. Физический электротон [Б71] - изменение значения мембранного потенциала, создаваемое пропусканием через данный участок мембраны электрического тока от внешнего (для данного участка мембраны) источника подпороговой силы. Это «пассивное» явление, определяемое внешним током и физическими свойствами покоящейся мембраны. Различают физический катэлектротон (рис. 2091921001), создаваемый выходящим током, и физический анэлектротон (рис. 2091921002), создаваемый входящим током. Физиологический электротон — это изменение возбудимости мембраны, создаваемое пропусканием через данный участок мембраны электрического тока от внешнего (для данного участка мембраны) источника подпороговой силы. Это «активное» явление, определяемое внешним током и физиологическими свойствами покоящейся мембраны. Различают физиологиский катэлектротон (рис. 2091921001), создаваемый выходящим током, и физиологический анэлектротон (рис. 2091921002), создаваемый входящим током. Физиологический электротон наблюдается в начале действия тока, при длительном действии он сменяется катодической депрессией и анодической экзальтацией.
Рис. 2091921001. Электрофизиологические феномены под катодом при действие постоянного подпорогового тока на возбудимые структуры. Элемент рис. 209192100.
Рис. 2091921002. Электрофизиологические феномены под анодом при действие постоянного подпорогового тока на возбудимые структуры. Элемент рис. 209192100.
Приставки кат- и ан- указывают на то, что такие токи и состояния возникают в области приложения к возбудимой структуре соответственно катода и анода. Ещё раз подчеркнём, что приставки кат‑ и ан‑ используются для случая внеклеточного расположения электродов.
Рис. 209201305. Выраженность катэлектротона и анэлектротона на разных участках нервного ствола вначале действия постоянного подпорогового тока.
Точнее эти изменения возбудимости показаны на рис. 2092013053 [++501+].
Рис.. Выраженность катодической депрессии и анодической экзальтации на разных участках нервного ствола при длительном действии постоянного подпорогового тока.
7. Замыкательно‑размыкательные законы (полярный закон) Э.Ф.В.Пфлюгера При раздражении нерва или мышцы постоянным током возбуждение возникает в момент замыкания постоянного тока только под катодом, а в момент размыкания — только под анодом. Эту закономерность открыл в 1859 г. Э.Пфлюгер. Как это было сделано? Умертвили участок нерва (рис. 209231300). При этом электротоническое проведение тока на поврежденном участке сохранилось, а возбудимость этого участка исчезла. Один из электродов установили на поврежденном участке, а второй — на неповрежденный.
Рис. 209231300. Полярный закон Э.Пфлюгера. Объяснение в тексте.
Если с неповрежденным участком соприкасается катод, возбуждение возникало в момент замыкания тока. Если же катод устанавливали на поврежденном участке, а анод — на неповрежденном, возбуждение возникает только при размыкании тока[Б72].
Рис.. Анодно-размыкательное возбуждение. КУД – критический уровень деполяризации, ПП – потенциал покоя, ПД – потенциал действия, АЭТ – анэлектротон, АЭ – анодическая экзальтация. Объяснение в тексте.
Рис.209251057
При более сильном токе происходят в принципе такие же процессы, но скорость нарастания деполяризации увеличивается и, следовательно, повышается частота генерирования потенциалов действия. Сравните рис.209251057 и рис.209251135.
Рис.209251135
При длительном действии деполяризующего стимула частота генерирования постепенно уменьшается (рис. 209251205). Это явление называется адаптацией [++501+C.48].
Рис. 209251205
Рис.209251235
Частотный оптимум и пессимум [V.G.75] ритмической стимуляции
Казалось бы, с увеличением частоты стимулов должна непрерывно увеличиваться частота ответов возбудимых структур, естественно до предела, определяемого лабильностью раздражаемой структуры. Однако на деле ситуация оказывается сложнее (рис. 209231607).
Рис. 209231607. Зависимость частоты ответов возбудимой структуры с рефрактерностью 5 мс (для стимула максимальной силы) от частоты стимуляции.
Для облегчения понимания введём конкретные числовые выражения. Условно определим интервалы временной продолжительности периодов рефрактерности модельной возбудимой структуры: 1. абсолютная рефрактерность ‑ 5 мс, 2. относительная рефрактерность – 5 мс.
Отсюда можно рассчитать частоту ритма раздражения, при котором импульсы раздражающего тока будут совпадать с той или иной фазой возбудимости.[++484+C.242]
Расчёт показывает, что при раздражении с частотой ритма менее 100 Гц все пороговые и сверхпороговые стимулы будут восприниматься раздражаемой структурой. При частоте стимуляции более 100 Гц число воспринятых стимулов в единицу времени будет зависеть от силы стимулов, их способности преодолеть порог раздражения при относительной рефрактерности (рис. 209231646 A).
Рис.209231646. Соответствие числа ответов (R) числу стимулов (S). Заштрихованные клеточки соответствуют абсолютной рефрактерности возбудимой структуры. Одна клеточка = 1 мс.
Рассмотрим случай, когда сила стимулов достаточная чтобы преодолеть любой период относительной рефрактерности, тогда препятствием для восприятия стимула будет только абсолютная рефрактерность. Т.е. рефрактерность будет равна 5 мс. Согласно нашей концептуальной модели раздражение с частотой чуть менее 200 Гц даст максимально возможную частоту ответов равную тем же 200 Гц (рис. 209231646 B).[V.G.76] как Как Следовательно, при раздражении с частотой ритма чуть более 200 Гц [V.G.77] многие импульсы тока будут действовать на ткань, когда она не способна отвечать на них. Ритм раздражения подвергается трансформации в более медленный ритм возбуждения. Будет неэффективен каждый 2‑й стимул (рис. 209231646 С). Частота ответов упадёт до 100 Гц. Дальнейший рост частоты стимуляции приводит к увеличению частоты возбуждений, но каждый второй стимул, а затем третий, четвёртый и т.д. стимул останется без ответа. На рис.209231607 представлена зависимость частоты ответов от частоты стимуляции.
Как видно из графика, максимальная частота ответов которую мы можем достичь при выбранных параметрах стимула и возбудимой структуры равна 200 Гц. Частота при которых достигается максимальная частота ответов названы Н.Е.Введенским оптимальной частотой. На графике мы видим, что в нашем примере оптимальными являются частоты кратные 200 Гц — 200, 400, 600, 800 и т.д. Сразу за оптимальными следуют пессимальные частоты, по Н.Е. Введенскому высокие частоты раздражения, вызывающие уменьшение ответа (201, 401, 601, 801 и т. д. Гц).
Как изменятся частотные оптимумы и пессимумы раздражения при снижении силы стимуляции? Очевидно, для этих стимулов возрастёт длительность рефрактерного периода. Как видно из графика (рис. 209231608), максимальная частота ответов, которую мы можем достичь при выбранных параметрах стимула и возбудимой структуры, равна 125 Гц. Оптимальными являются частоты кратные 125 Гц.
Рис. 209231608. Зависимость частоты ответов возбудимой структуры с рефрактерностью 8 мс (для стимула средней силы) от частоты стимуляции.
При использовании пороговой стимулов рефрактерность возбудимой структуры будет равна АРП+ОРП, в нашем примере 10 мс. Как видно из графика (рис. 209231609), в этом случае максимальная частота ответов, которая достижима при выбранных параметрах стимула и возбудимой структуры равна 100 Гц. И как мы уже отметили, именно этой частоте кратны оптимальные частоты раздражения.
Рис. 209231608. Зависимость частоты ответов возбудимой структуры с рефрактерностью 8 мс (для стимула средней силы) от частоты стимулции.
Можно сделать вывод, что если выполняются законы раздражения (силы, времени, градиента) для одиночного стимула, то оптимум при серийных раздражениях этим стимулом будет не больше 1/АРП и не меньше 1/(АРП+ОРП). Т.е. лабильность определяется длительностью рефрактерных периодов. Лабильность неодинакова не только у разных тканей, но и у разных структурных единиц одной и той же ткани. Более того, даже у клетки лабильность непостоянна и определяется ее функциональным состоянием. Не следует путать понятия «усталость», «пессимальное торможение» и пессимальная частота раздражения. Если мы наблюдаем пессимальную частоту раздражения, стоит нам незначительно уменьшить частоту стимуляции и мы отметим значительный рост частоты возбуждений.
Рис. 209251331. Зависимость частоты ответов возбудимой структуры от частоты стимулов..
При усталости и пессимальном торможении этого не произойдет. Причину мы выясним в дальнейшем. Парабиоз Н.Е.Введенского Экспериментальные факты, составляющие основу учения о парабиозе, Н.Е.Введенский (1901) изложил в своем классическом труде «Возбуждение, торможение и наркоз». Опыты проводились на нервно-мышечном препарате. Схема опыта показана на рис. 2092313240 и 209231324.
Нервно-мышечный препарат помещался во влажную камеру, а на его нерв накладывались три пары электродов: 1. для нанесения раздражения (стимуляции) 2. для отведения биотоков до участка, на который предполагалось воздействовать химическим веществом. 3. для отведения биотоков после участка, на который предполагалось воздействовать химическим веществом.
Кроме этого, в опытах регистрировались сокращение мышцы и потенциал нерва между интактным и альтерированным участками. О частоте следования импульсов после альтерированного участка можно было судить по наличию, характеру и амплитуде тетанического сокращения икроножной мышцы. Но к этому мы вернёмся изучив физиологию мышечного сокращения (лекция 5). Если же участок между раздражающими электродами и мышцей подвергнуть действию наркотических веществ и продолжать раздражать нерв, то ответ на раздражение через некоторое время исчезает[V.G.78].
Рис. 209231324. Схема опыта
Н.Е.Введенский, исследуя в подобных условиях действие наркотиков и
|
||||||
|
|
Последнее изменение этой страницы: 2016-06-29; просмотров: 1450; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.102 (0.016 с.) |
















 [V.G.70]
[V.G.70]