Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Гормональный и физиологический контроль линьки у птиц.Содержание книги
Поиск на нашем сайте
Смена оперения происходит периодически, в определенные сезоны года. Линька всегда идет постепенно. Линька зависит от многих факторов. Существует несколько нарядов птицы: эмбриональный, гнездовой, послегнездовой. Линьки бывают сезонные, полная годовая линька, предбрачная линька. При линьке меняются не только перья, но и другие покровные образования. В период линьки происходят изменения в организме: Содержание жира в теле в это время резко снижается до минимального в годовом цикле уровня. Напротив, сырая нежирная масса тела (масса тела за вычетом жира) в этот период более высока, чем до линьки и после нее, что в первую очередь определяется увеличением количества воды, достигающего наиболее высокого уровня в годовом цикле. При этом на протяжении линьки количество воды в теле изменяется параллельно интенсивности смены оперения. Перед линькой увеличивается количество белков — они составляют около двух третей сухой нежирной массы тела. В начальном периоде линьки количество белков в теле снижается, во второй ее половине происходит восстановление их содержания до исходного уровня. Энергетическая стоимость линьки, определенная у 15 видов воробьинообразных птиц, составляет 540—2595 кДж. Гормональный контроль линьки: - гормон щитовидной железы – обеспечивает необходимый для перообразования обмен веществ, образование окраски развивающегося пера, дифференцировка перьевого зачатка; - гормон семенников: перья брачного наряда у самцов; - женские половые гормоны: окраска самок. - половые стероидные гормоны: торможение линьки у видов, у которых она не связана с размножением: домашние голуби-куры-канарейки. - гормоны гипофиза: определяют уровень половых стероидов, обеспечивают периодичность линьки. -пролактин+АКТГ замедляет и ингибирует линьку (голуби, зяблик) На линьку влияет и фотопериод. Это внешний фактор, синхронизирующий связанные с линькой физиологические изменения с сезонными явлениями. 18. ПОСЛЕГНЕЗДОВОЙ ПЕРИОД В ЖИЗНЕННОМ ЦИКЛЕ ПТИЦ. Послегнездовые кочевки начинаются с уходом слетков из гнезда и заканчиваются у оседлых птиц занятием территории будущего гнездования, у перелетных — началом активного пролета в направлении мест зимовки. После вылета связь молодых птиц с территорией постепенно усложняется: до распадения выводка ее осуществляют взрослые птицы, собирающие корм для следующих за ними слетков, т. е. связь носит преимущественно пассивный характер. Распадение семейной группировки способствует установлению активной связи с территорией, особь оценивает свое положение в пространстве в соответствии с наследственной программой и опытом, приобретенным в период следования за взрослыми птицами. Важным событием этого периода является послегнездовая дисперсия - расселение молодых птиц. Массовое кольцевание позволило получить сведения о радиусе дисперсионного разлета, о поведении этих птиц в период дисперсии.
Для многих видов воробьиных послегнездовой период — это время линьки, в процессе которой существенно возрастают потребности в корме. Поскольку в ходе выкармливания птенцов определенная часть корма на гнездовом участке выедается, птицы в послегнездовое время активно перемещаются по территории в поисках участков и биотопов, мало посещавшихся в гнездовое время. Послегнездовые кочевки выводков Поведение слетков в первые дни после оставления гнезда описано неоднократно и не отличается большим разнообразием. В течение 5 — 7 дней после вылета слетки находятся в непосредственной близости от места вылупления, связи между собой не поддерживают, за взрослыми птицами не перемещаются или делают это неактивно. Затем активность молодых резко возрастает, происходит объединение слетков в одну или две группы (по числу взрослых птиц, докармливающих выводок); может произойти смена биотопов. О моделях послегнездовых кочевок Молодые птицы в период послегнездовых кочевок находятся под влиянием двух основных факторов — внутренней программы развития и внешних условий. Программа определяет сроки распадения выводка, длительность нахождения особи в районе вылупления, возможность участия в дисперсионном разлете, сроки начала дисперсии, длительность периода дисперсии и, вероятно, расстояние, преодолеваемое птицей при дисперсионном разлете. Внешние факторы, прежде всего трофические, определяют местное направление и биотопическую предпочтительность кочевок, возможно, корректируют сроки распадения выводков. Влияют внешние факторы и на длительность нахождения особи в контрольном районе: в богатых кормом биотопах птицы находятся дольше. Немаловажное значение имеют климатические условия: в неблагоприятную погоду птицы малоподвижны. На поведение птиц влияет физиологическое состояние особей, прежде всего — участие в линьке и полнота этой линьки. Практически у всех изученных видов в период активной линьки подвижность птиц снижалась.
Разная доля участвующих в дисперсии птиц, разная длительность периодов покоя и разлета создают характерную для вида схему кочевок. Повидовые схемы можно объединить в несколько типов-моделей после гнездовых кочевок: Модель таловки для тех, у кого нет пост ювенильной линьки, в местах гнездования, дальние мигранты. Дисперсия у первого выводка, у 2го – сразу миграции. Для модели веснички характерно кратковременное пребывание в районе рождения; участие в дисперсионном разлете преобладающей части местных молодых птиц; наличие после дисперсионного периода относительного покоя. Модель варакушки применима к видам с растянутым периодом покидания гнездового района — варакушке, а также, видимо; белобровику и рябиннику, сибирской завирушке. Несколько периодов ухода птиц из гнездового района, перемещения дисперсирующих и кочующих птиц образуют ряд последовательных волн численности, чаще 4 —6, из которых каждая последующая обычно ниже предыдущей. Модель чечетки характеризуется быстрой утратой связи молодой птицы с районом рождения, включением в стаю, где молодые особи, вероятно, занимают зависимое положение и не определяют продолжительность нахождения стаи в данном районе. Направление движения птиц и весь характер после гнездовых кочевок непредсказуемы. Направление после гнездовых кочевок Дисперсионный разлет не должен иметь выраженного направления движения, но поскольку перемещения птиц после распада выводков осуществляются по богатым кормом биотопам, всегда есть местный вектор движения — по долинам ручьев, рек, горным долинам, по морскому побережью. Поведение взрослых птиц после распадения выводка не отличается разнообразием. У большинства видов начинается интенсивная линька маховых, они частично утрачивают способность к полету, держатся скрытно и, видимо, мало перемещаются. 19. ПОЛЕТ ПТИЦ. ТИПЫ СТРОЕНИЯ КРЫЛА. АЭРОДИНАМИКА ПОЛЁТА. Строение крыла приспособлено к образу жизни данной птицы, будь то ласточка, парящий кондор или передвигающийся по земле фазан. Наиболее обычные типы крыла: 1. Эллиптическое крыло. Птицы, живущие в лесах и на земле, такие как куриные, голуби, дятлы и многие воробьиные, имеют короткие широкие крылья с множеством щелей (изменяемых промежутков между маховыми перьями первого порядка). Такое строение обеспечивает высокую маневренность и быстрый взлет. 2. Высокоскоростное крыло. Птицы, кормящиеся подобно стрижам в воздухе или совершающие длительные миграции, как, например, крачки, имеют длинные, относительно узкие крылья. Такое крыло лучше приспособлено к быстрому равномерному полету, чем к скоростному взлету и маневрированию в ограниченном пространстве.
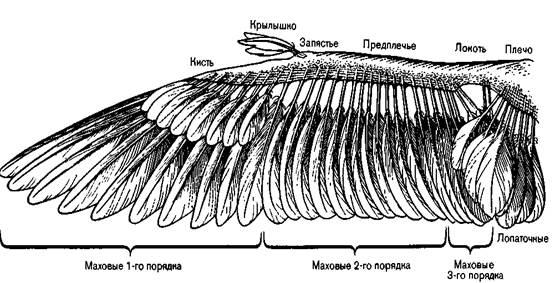
3. Крыло с высоким отношением длины к ширине. Очень длинные, узкие крылья парящих морских птиц, таких как альбатросы и буревестники, приспособлены к высокоскоростному планированию при сильных устойчивых ветрах. 4. Щелевое крыло, создающее большую подъемную силу. Птицы, парящие над сушей, такие как кондоры, грифы, орлы и совы, имеют длинные широкие крылья с множеством щелей. Такое строение крыла позволяет сочетать маневренность с плавным скольжением, дает возможность птице кружить в небольших восходящих потоках теплого воздуха, образующихся над землей. Колибри имеет очень своеобразный полет: они могут подниматься вертикально вверх, зависать в воздухе и летать назад. Крылья колибри, когда она зависает описывают восьмерку в воздухе. Чем меньше птица, тем больше отношение площади крыла к весу. Птица при полете создает разницу давлений у крыла. При полете крылья опускаются, одновременно смещаясь вперед. Воздух плавно обтекает тело птицы. Маховые 1го порядка – асимметричные, пропеллирующий эффект, могут вращаться вокруг своей оси. Локтевая+лучевая кости – симметричные перья. Маховые 2го порядка. Маховые 3го порядка – полетная поверхность. Абсолютная симметричность. Кроющие перья – перья сверху. Крылышко – щель при взлете и посадке, аэродинамика торможения. Хвост – рулевые перья. Крепятся к пигостилю. Самые крайние перья – асимметричные, край у перьев круглый. Морские птицы могут парить против господствующего направления ветра, используя восходящую тягу на стыке 2х воздушных цилиндров вращающихся в противоположных направлениях. Есть птицы использующие встречный ветер (олуши). Есть птицы, которые парят над землей, например крупные горные падальщики.
|
|||||||
|
|
Последнее изменение этой страницы: 2016-04-26; просмотров: 1157; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.22.242.43 (0.009 с.) |