Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Сравнение количества жидкости в организмах ребенка и взрослогоСодержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте Общее количество жидкости на 1 кг массы тела у детей больше, чем у взрослых, однако водный обмен у детей протекает более интенсивно. Так, у взрослых вся вода в организме обновляется примерно каждый месяц, а внеклеточное водное пространство – каждую неделю. У грудного же ребенка время пребывания молекулы воды в организме составляет всего 3–5 дней. В отличие от взрослых, у детей раннего возраста отмечается большая проницаемость клеточных мембран, а фиксация жидкости в клетке и межклеточных структурах более слабая. Величину пространств, занимаемых жидкостью, можно определить с помощью специальных методов с применением таких веществ, которые распределяются равномерно или во всех жидкостях организма, или только во внеклеточной жидкости (хлоридное пространство), или только в плазме крови. Либо можно определить лабильную, не связанную с кислыми мукополисахаридами воду (инулиновое пространство). У ребенка объемы хлоридного и инулинового пространств почти одинаковы: 41,2 и 40 % от массы тела. У взрослого они соответственно равны 26,4 и 16 %. Это свидетельствует о большей скорости протекания водного обмена у детей. С помощью изотопов можно определить общее содержание натрия, калия и хлора в организме. Классификация жидкостей Хотя жидкости в организме представляют собой единое целое, химический состав их различен. Принято различать внеклеточную и внутриклеточную жидкости, которые разделены мембранами клеток. При зваболеваниях может появляться третье водное пространство – в полостях тела: брюшной, плевральной и т. д. Внеклеточная или экстрацеллюлярная жидкость Объем внеклеточной жидкости составляет 20–25 % от массы тела. Состоит из: · жидкой части или плазмы крови (5 % от массы тела), которая образует транспортирующую систему, питающую клетки и уносящую продукты обмена веществ; · тканевой или интерстициальной жидкости (15 % от массы тела); · трансцеллюлярной жидкости (1–3 % от массы тела), которая, в свою очередь, состоит из секретов желудочно-кишечного тракта и спинномозговой, внутриглазной, плевральной, перитонеальной, синовиальной жидкостей. Объем трансцеллюлярной жидкости значительно увеличивается при воспалительных заболеваниях кишечника (диарее, кишечной непроходимости, плеврите и др.). Благодаря сходному химическому составу плазма и интерстициальная жидкость составляют функциональное единство. Внеклеточная жидкость также подразделяется на: · мобильную, свободную часть плазмы, располагающуюся между клетками и капиллярами; · менее мобильную, находящуюся в соединительной ткани. Внеклеточная жидкость связана с внешней средой через желудочно-кишечный тракт, легкие, кожу и почки. Жизненно важной функцией внеклеточной жидкости является поддержание нормального количества плазмы крови и тем самым обеспечение кровообращения. При недостатке воды в организме вода из тканей направляется в плазму крови, при избытке наоборот. Накопление или потеря электролитов также отражаются на ионном соотношении как плазмы, так и всей внеклеточной жидкости. Внеклеточная жидкость является внутренней средой, в которой живут клетки. Следовательно, она должна обеспечивать оптимальные условия для их жизнедеятельности. Уменьшение с возрастом внеклеточной жидкости в основном обусловлено увеличением роста клеток и уменьшением скорости роста коллагена по отношению к мышечной ткани. Внутриклеточная или интрацеллюлярная жидкость Объем внутриклеточной жидкости составляет в среднем около 40 % от массы тела. Она представляет собой сумму жидкостей в клетках, имеющих разные локализацию, функцию, состав. Водный баланс складывается из трёх процессов: Изменения или нарушения водного обмена обозначаются как положительный (накопление в организме избытка воды) или отрицательный (дефицит в организме воды) баланс. Суточный баланс воды в организме взрослого человека
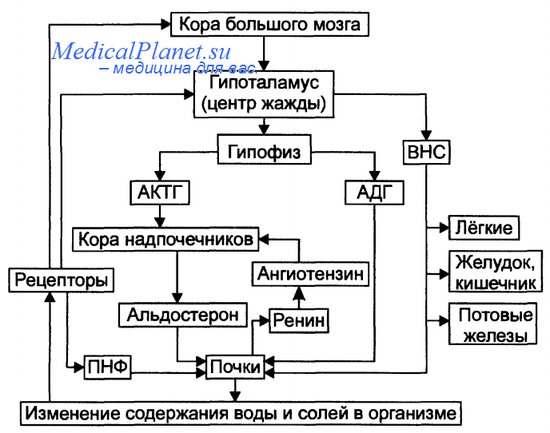
Регуляция обмена воды Система регуляции обмена воды имеет сложную структуру. Адаптивная цель этой системы — поддержание оптимального объёма жидкости в организме. При воздействии патогенных факторов и/или отклонении содержания жидкости и солей в организме эта система устраняет сдвиги или способствует уменьшению их степени. Функция системы регуляции водного обмена тесно связана с системами контроля солевого обмена и осмотического давления. Система регуляции обмена воды в организме включает центральное, афферентное и эфферентное звенья.
Система регуляции водного обмена организма. ВНС — вегетативная нервная система; ПНФ — предсердныи натрийуретический фактор (атриопептин); рецепторы — чувствительные нервные окончания. Афферентная импульсация от рецепторов различного тип а (хемо-, осмо-. баро-, терморецепторов, возможно, и некоторых других) поступает к нейронам гипоталамуса. Наиболее важное значение при этом имеют: Регуляторные стимулы от нейронов центра жажды (нервные и гуморальные) адресуются эффекторным структурам. • Эфферентное звено системы регуляции водного обмена включает почки, потовые железы, кишечник, лёгкие. Эти органы в большей (почки) или в меньшей (например, лёгкие) мере обеспечивают устранение отклонений содержания воды, а также солей в организме. Важными регуляторами главного механизма изменения объёма воды в организме — экскреторной функции почек — являются антидиуретический гормон (АДГ), система «ренин—ангиотензин—альдостерон», предсердныи натрийуретический фактор (атриопептин), катехоламины, Пг, минералокортикоиды. При воздействии патогенных факторов и/или отклонении содержания жидкости в организме система регуляции водного обмена, как правило, устраняет эти отклонения или обеспечивает уменьшение их степени. Если же эффективность этой системы недостаточна, развиваются различные варианты нарушений водного обмена. Антидиуретический гормон (АДГ). Вазопрессин Каждый день почки фильтруют неодинаковое количество шлаков и воды. Они реагируют на изменения потребностей организма. Например, при повышенной температуре организм с потом теряет больше воды. Это может привести к обезвоживанию. Чтобы компенсировать процесс потоотделения, почки реабсорбируют из канальцевой жидкости больше воды, чем обычно. В результате мочи выделяется меньше, и она становится более концентрированной и Стандартной диагностической процедурой является удаление воды из мочи и анализ осадка. Каким образом обеспечивается контроль? Это сложное взаимодействие нервов и гормонов, которые ежеминутно реагируют на перемены в организме. Представьте, что вы активно двигаетесь в жаркий день. Вы начинаете потеть, уровень воды в организме и, следовательно, в крови падает. По мере того, как кровь циркулирует через мозг, она активирует специальные сенсоры (осморецепторы) в гипоталамусе, расположенном на стволе головного мозга. Осморецепторы посылают по нервам сообщения в гипофиз — орган размером с горошину у основания мозга. Эти сообщения стимулируют гипофиз к высвобождению АДГ (антидиуретического гормона, вазопрессина). Он поступает в кровь и переносится к почкам. В почках АДГ воздействует на внутреннюю выстилку дистальных извитых канальцев и собирательных трубок. Его воздействие повышает проницаемость поверхности. Под влиянием АДГ большее количество воды переходит из жидкости, находящейся в канальцах и трубочках, обратно в кровь расположенных вокруг них капилляров. Моча становится более концентрированной, а кровь более жидкой за счёт дополнительной воды. Конечным результатом является меньшее количество мочи и сохранение воды в организме. Противоположные процессы наблюдаются, если в течение короткого промежутка времени выпить большое количество жидкости. Количество воды в крови моментально увеличивается. Осморецепторы мозга не раздражаются, они перестают посылать в гипофиз сигналы о необходимости высвободить АДГ. Соответственно, содержание АДГ, переносимого кровью к почкам, снижается. Внутренняя выстилка канальцев и трубочек начинает пропускать через себя меньше воды. В моче остаётся больше жидкости. Результатом является более обильная и менее концентрированная моча. Осморецепторы (от осмос и рецепторы), концевые образования чувствительных нервов, реагирующие на изменение концентрации осмотически активных веществ, т. е. осмотического давления (ОД), в окружающей их жидкости. О. имеются в различных органах и тканях животных (печень, поджелудочная железа и др.). Импульсы от О. поступают в отделы центральной нервной системы, участвующие в регуляции водно-солевого обмена организма (см. также Осморегуляция). Функция осморецепции присуща нервным клеткам т. н. супраоптических ядер гипоталамуса, которые воспринимают даже небольшие колебания ОД плазмы крови (1—2%) и при повышении ОД вырабатывают гормон вазопрессин, накапливающийся в гипофизе. Выделяясь в кровь, этот гормон, обладающий антидиуретическим действием, изменяет проницаемость стенки почечных канальцев, что приводит к уменьшению объёма выделяемой мочи. Осморегуляция (от осмос и лат. regulo — направляю), совокупность физико-химических и физиологических процессов, обеспечивающих относительное постоянство осмотического давления (ОД) жидкостей внутренней среды (межклеточных жидкостей, лимфы, крови) у гомойосмотических животных. Механизмы О. обнаруживаются у организмов, обитающих в средах с различной концентрацией осмотически активных веществ, главным образом солей, и при разном уровне потребления как этих веществ, так и воды. О. присуща всем пресноводным и наземным животным; среди морских организмов О. обладают все позвоночные (кроме миксин), некоторые ракообразные. В основе физиологических механизмов О. лежит соответствующий рефлекс: изменение ОД крови или межклеточной жидкости воспринимается осморецепторами; возникающие в них импульсы передаются в нервные центры, которые регулируют потребление воды и солей и их выделение осморегулирующими органами — нефридиями, почками, солевыми железами и др. Механизм О. может быть гиперосмотическим, когда ОД внутренней среды больше, чем в жидкости внешней среды, и гипоосмотическим, когда внутреннее ОД меньше внешнего. В первом случае избыток воды выделяется животным главным образом через почки (растением — преимущественно через устьица); во втором — пассивную потерю воды через покровы тела организм восполняет, потребляя богатую солями воду и выделяя избыток солей в основном через солевые железы. Все пресноводные животные и морские хрящевые рыбы характеризуются гиперосмотических регуляцией. У акул и скатов это обусловлено высокой концентрацией в крови мочевины. В тело этих животных через проницаемые для воды участки покровов по осмотическому градиенту поступает вода. Избыток её у всех животных выделяется почками или их аналогами (сократительные вакуоли простейших, нефридии), а соли всасываются из пресной воды клетками жаберного аппарата или кожи (у земноводных). Гипоосмотическая регуляция характерна для морских костистых рыб, морских пресмыкающихся и некоторых других. Эти животные теряют воду через покровы и с мочой. Для компенсации потерь они пьют морскую воду, опресняемую клетками солевых желёз и др. органами, выделяющими концентрированные растворы солей. У млекопитающих основной орган О. — почки, способные выделять гипотоническую мочу при избытке воды и осмотически концентрированную — при её дефиците. Проходные рыбы (например, лососи) и некоторые ракообразные обладают обоими механизмами О. и могут жить как в пресной, так и в морской воде. Важнейшими барорецепторными зонами (рецепторов растяжения, воспринимающих АД) являются области дуги аорты и каротидного синуса (рис. 18-27). Если сосудистую стенку растягивать под действием постоянного давления, то импульсация в барорецепторах будет непрерывной, причем кривая зависимости частоты этой импульсации от давления имеет почти S-образный характер. Импульсы от них по синокаротидному нерву (нерву Геринга, ветви языкоглоточного нерва) поступают в продолговатый мозг. Барорецепторы реагируют на колебания артериального давления во время сердечного цикла ритмичными залпами разрядов, частота которых изменяется тем больше, чем выше амплитуда и скорость нарастания волны давления. Поэтому барорецепторы передают информацию не только о среднем артериальном давлении, но также об амплитуде колебаний давления и крутизне его нарастания, а следовательно, и о ритме сердца. Афферентные импульсы от барорецепторов поступают к кардиоингибиторному центру и сосудодвигательному центру, расположенным в продолговатом мозге, а также в другие отделы ЦНС. От рецепторов растяжения, расположенных в артериях и венах большого и малого крута кровообращения (барорецепторов и волюморецепторов), в ствол мозга постоянно поступает информация об артериальном и венозном давлении. На основании этой информации изменяется симпатический тонус, благодаря чему поддерживаются АД и кровоток в жизненно важных органах (рис. 70.3). При повышении АД увеличивается импульсация от барорецепторов каротидного синуса и дуги аорты. Эта импульсация поступает в ядро одиночного пути продолговатого мозга, а затем - в стволовые симпатические центры, оказывая на них тормозное влияние (рис. 70.3). Благодаря этому так называемому барорефлексу повышение АД по механизму отрицательной обратной связи приводит к снижению симпатического тонуса. В этом торможении участвует стволовой норадренергический путь, связанный с ядром одиночного пути. Нейроны этого пути возбуждаются центральными альфа2-адреностимуляторами, и именно на них действуют некоторые гипотензивные средства (например, клонидин), усиливающие барорефлекторную депрессорную реакцию. Когда же АД падает, снижение импульсации от барорецепторов приводит к уменьшению тормозных влияний на стволовые симпатические центры, что сопровождается повышением симпатического тонуса и ростом АД. ВОЛЮМОРЕЦЕПТОРЫ (от франц. volume - объем и рецепторы), чувствительные нервные окончания в различных органах и кровеносных сосудах, реагирующие на растяжение их стенок. Обмен воды и электролитов в организме имеет сложную нервно-гуморальную регуляцию. Наиболее подвержены регуляторным воздействиям вода и тесно связанный с нею в метаболизме натрий. Сложная рефлекторная цепь регуляции водно-электролитного обмена начинается с четырех рецепторов, которые сигнализируют об изменении количества воды в организме. Во-первых, это рецепторы слизистой рта, подсыхание которой вызывает чувство жажды. Однако это ощущение проходит при смачивании слизистой, хотя вода в организм при этом не поступает. Поэтому данный вид жажды называется ложной жаждой. Во-вторых, сигналы о необходимости восполнить запасы воды в организме или прекратить ее потребление идут от барорецепторов слизистой желудка. Раздувание желудка ведет к исчезновению чувства жажды, а спадение его стенок - наоборот, к возникновению. Поскольку жажда, возникающая в этом случае, связана не с изменением содержания воды в организме, а с изменением тонуса желудочной стенки, она также ложная. Третью группу нервных окончаний, принимающих участие в регуляции водно-электролитного обмена, составляют осморецепторы тканей, которые сигнализируют об изменениях осмотического давления в тканях. Чувство жажды при раздражении осморецепторов - это вид жажды истинный. И наконец, четвертая группа рецепторов - волюм-рецепторы сосудистого русла, реагирующие на изменение объема циркулирующей в сосудистой системе крови. В регуляции водно-солевого обмена принимают участие и дистантные рецепторы (зрительный, слуховой), обеспечивающие условнорефлекторный компонент регуляции. Импульсы со всех указанных групп рецепторов идут в гипоталамус, где расположен центр водорегуляции. Отсюда поступают "команды" на эффекторы, выводящие воду из организма. Регулятором водно-солевого обмена являются гормоны коры надпочечников (альдостерон) и задней доли гипофиза (антидиуретический). Ангиотензины (Angiotensins): физиологически активные пептиды широкого спектра действия. Обнаружены практически во всех тканях организма, включая структуры центральной нервной системы. Пептиды группы ангиотензина участвуют в регуляции не только уровня артериального давления и сопряженных процессов почечной фильтрации и водно-солевого обмена, но также и в репродуктивной функции, многих процессах генерализованного характера (стресс, алкогольная мотивация, агрессивное поведение), процессах ноотропного ряда. Ангиотензин участвует в синтезе или рилизинге ряда других физиологически активных соединений - гормонов, низкомолекулярных медиаторов (катехоламины, серотонин), чем в значительной мере объясняется широкий спектр физиологических функций этих субстанций. Исследования последнего времени уделяют внимание молекулярным аспектам рецепторов ангиотензина II. Обзоры: Jin M. et al., 1989, Eberhardt R. et al., 1993, Schunkert H. et al., 1993, Kang P. et al., 1994, Wright J. et al., 1994. Ангиотензины - пептидные гормоны, образующиеся в крови и периферических тканях при действии специфических протеаз (в частности ренина) из крупномолекулярного белкового предшественника - прогормона (ангиотензиногена), секретируемого печенью. Первичный продукт протеолиза неактивного ангиотензиногена - декапептид ангиотензин I. Из ангиотензина I за счет отщепления двух аминокислотных остатков с С-конца его молекулы образуется основной эффекторный пептид - ангиотензин II под влиянием ангиотензин- превращающего фермента (дипептидил карбоксипептидазы, К.Ф.3.4.15.1). Этот фермент также участвует в инактивации брадикинина - физиологического антипода ангиотензина II и других пептидов. Ингибиторы ангиотензин-превращающего фермента нашли широкое распространение в клинической практике сердечно- сосудистых патологий. Ангиотензины обладают выраженными вазопрессорным действием, а также являются специфическими регуляторами биосинтеза альдостерона корой надпочечников. При этом ангиотензин II приблизительно на порядок активнее ангиотензина I. Очевидно, два С-концевых аминокислотных остатка декапеп- тида тормозят его биологическую активность. Проведенный функционально-структурный анализ молекулы ангиотензина II показал, что роль актона в ней, по-видимому, выполняют 1-3-трипептид, расположенный на N-конце, и 8-й (С-концевой) аминокислотный остаток. Роль же рецептор-связываемого локуса выполняет средний 3-6-тетрапептидный фрагмент молекулы. В 70х годах обнаружен ангиотензин III, обладающий повышенным сродством к рецепторам коры надпочечников. Ангиотензин III - гептапептид, образующийся из ангиотензина II путем отцепления от него N-концевого аспарагилового остатка. Ангиотензиновые рецепторы - G-белоксопряжённые рецепторы, лигандами которых является ангиотензин, гормон ренин-ангиотензиновой системы, и его производные. У человека описано четыре AT-рецептора. Антагонисты рецепторов ангиотензина II применяются в качестве средств, снижающих давление.
|
||
|
|
Последнее изменение этой страницы: 2016-04-26; просмотров: 697; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.141 (0.01 с.) |