Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Причины хромосомных аберрацийСодержание книги
Поиск на нашем сайте
Основной предпосылкой для возникновения хромосомных перестроек является появление в клетке двунитевых разрывов ДНК, то есть разрывов обеих нитей спирали ДНК в пределах нескольких пар оснований. Двунитевые разрывы ДНК возникают в клетке спонтанно или под действием различных мутагенных факторов: физической (ионизирующее излучение), химической или биологической (транспозоны, вирусы) природы. Двунитевые разрывы ДНК возникают запрограммированно во время профазы I мейоза, а также при созревании Т- и B-лимфоцитов во время специфической соматической V(D)J рекомбинации. Нарушения и ошибки процесса воссоединения двунитевых разрывов ДНК приводят к появлению хромосомных перестроек[3]. Генные мутации могут происходить на уровне кодирующих или регуляторных последовательностей структурных генов: в первом случае, изменяется структура (качество синтезированного полипептида), а во втором — изменяется скорость синтеза (количество) конечного продукта. В результате генных мутаций возникают альтернативные формы гена, называемые аллелями.
Механизмы генных мутаций: - нарушения последовательности нуклеотидов - замены, инверсии, делеции, инсерции (вставки) нуклеотидов; - внутригенные рекомбинации и неравный кроссинговер; реверсии; - дупликации и гипердупликации. Наиболее часто встречаются замены нуклеотидов, которые относятся к точечным мутациям и бывают двух типов: - трансверсии - замена пуринового основания на пиримидиновое и наоборот; - транзиции - замена одного пуринового основания на другой пуриновое или одного пиримидинового основания на другой пиримидиновое.
Замена приводит к изменению только одного кодона (смыслового или стоп-кодона). Замена смыслового кодона может приводить к: - замене аминокислоты, как следствию замены кодона - misens-мутация; - остановке синтеза полипептида, если в результате замены появился стоп-кодон (UAA,UAGşiUGA)- nonsens-мутация; - сохранению исходной (нормальной) структуры полипептида, если в результате замены образовался кодон-синоним, кодирующий ту же аминокислоту - samesens- мутация.
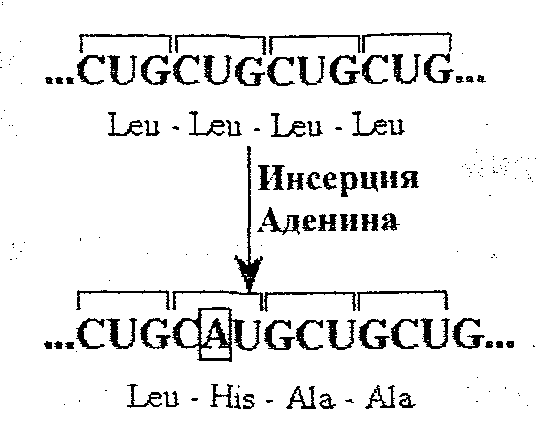
Замена иногда может возникать в стоп-кодоне, и тогда UAAилиUAGможет стать САА илиCAG, которые кодируют глутамин. В этом случае синтез полипептида продолжается до следующего стоп-кодона. Примером такого "удлинения цепи" может быть анормальный гемоглобинHb-HbCS(HbConstantSpring), который состоит из 172-х аминокислот вместо 141, а дополнительная последовательность действительно начинается глутамином. Замены могут происходить в разных кодонах, приводя к замене двух и более аминокислот в одной и той же полипептидной цепи. Например: HbC-Hariem= 2alfa2beta6Glu→Val; 73 Asp→Asn. Инверсия приводит к изменению последовательности нуклеотидов в кодоне и обратному чтению кодона. Последствия инверсии сходны с таковыми в случае замен. Делеция - это выпадение одной или нескольких пар нуклеотидов в молекуле ДНК. Последствия зависят от количества вовлеченных в делецию пар нуклеотидов. Если отсутствует одна пара нуклеотидов, то это приводит к сдвигу рамки чтения (мутация "frameshift "); а в синтезированном полипептиде все аминокислоты после делеции будут изменены (например,HbWayne). Если количество делетированных нуклеотидов превышает 3, в полипептидной цепи будет отсутствовать одна или более аминокислот (например, в HbGun-Hillотсутствуют 5 аминокислот в цепи р: 91-95). Иногда происходит делеция целого гена. При альфа-талассемии не образуются цепи а, т.к. данный ген отсутствует в геноме, и в этом случае образуются другие типы цепей:HbН=4betaилиHbBart=4gama. Инсерция, или вставка, представляет собой появление нового нуклеотида в последовательности гена, что приводит к сдвигу рамки чтения и синтезу измененного полипептида.
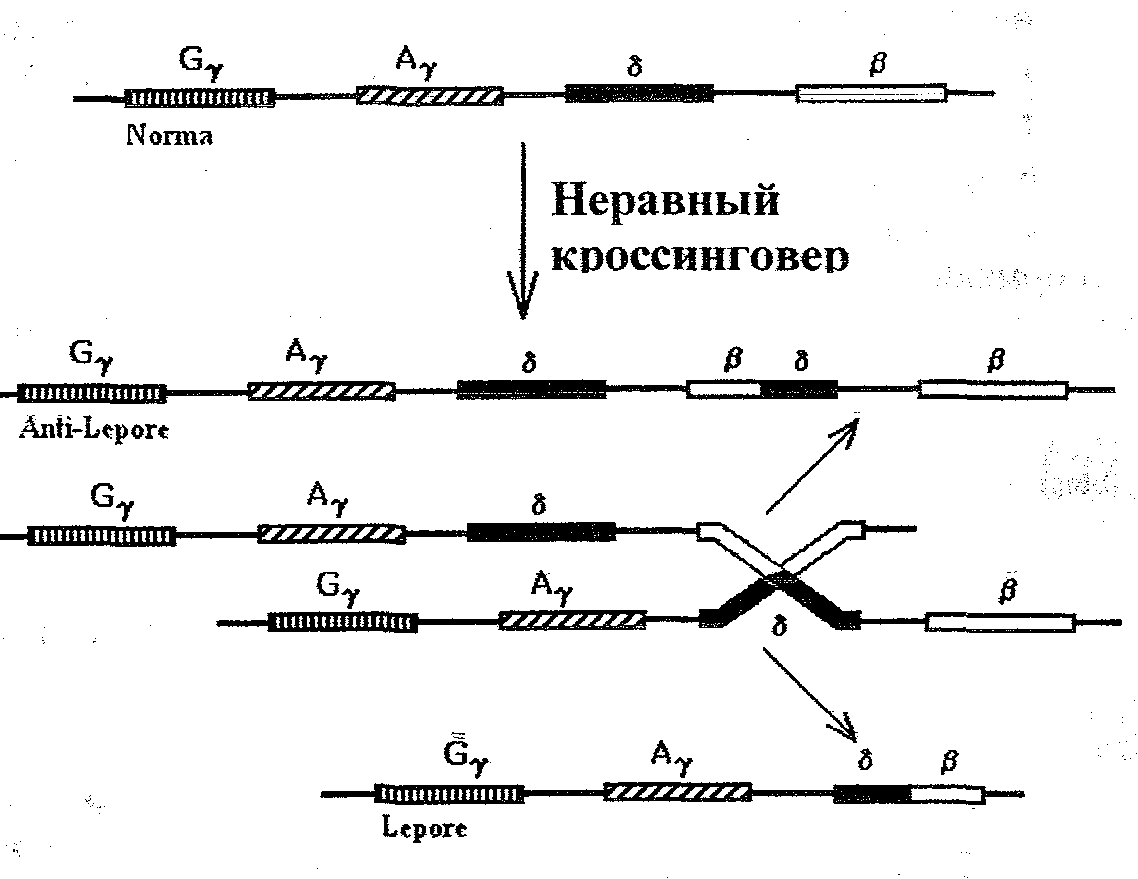
Неравный кроссинговер является следствием неправильной конъюгации гомологичных хромосом, в результате чего происходит перестановка нуклеотидных последовательностей ДНК и изменяется структура полипептида (например,HbLepbfe).
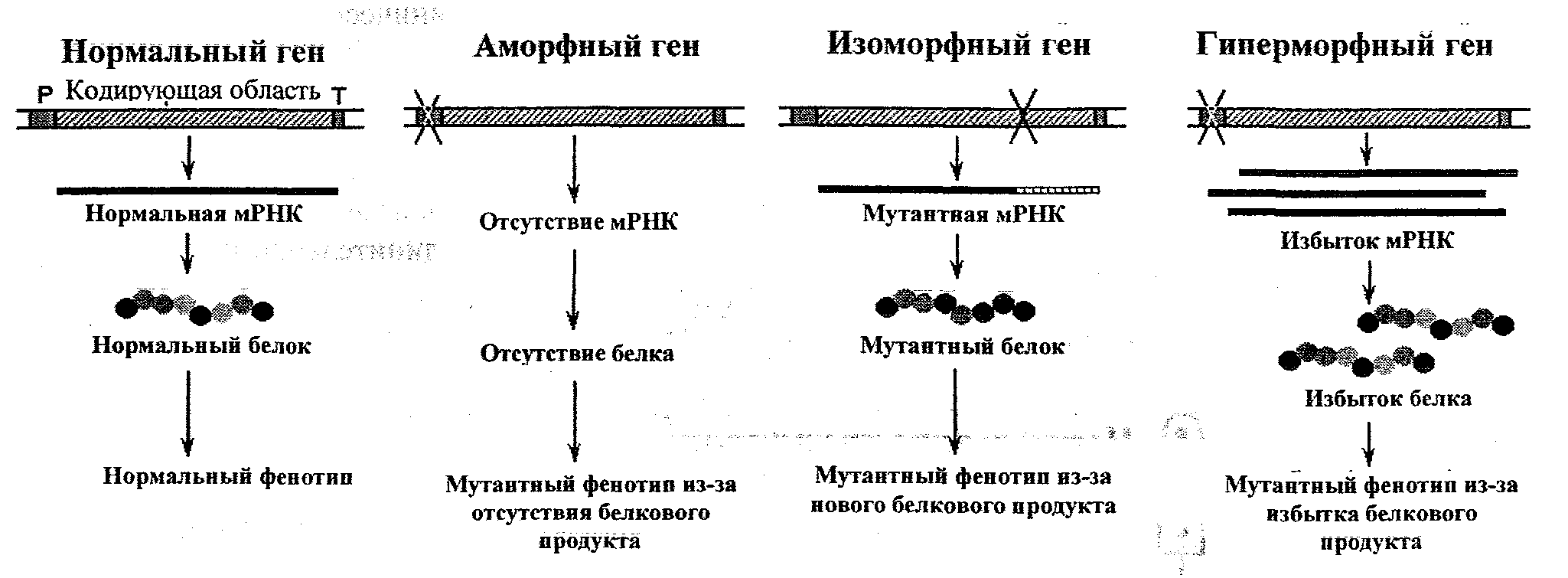
Реверсия (обратная мутация) - это изменение в мутантном гене, приводящее к возврату его в исходное нормальное состояние. Истинная реверсия трансформирует мутантный кодон в нормальный, а супрессивная реверсия индуцирует другую мутацию, которая отличается по положению, но имеет сходный эффект. Например Hb.Harlemсвязана с мутацией в цепи β6Glu-Val, как иHbS, но действие ее в виде образования эритроцитов анормальной формы аннулируется в результате обратной мутации в цепи β73Аsр-Аsn. Аналогичная ситуация в случаеHbMemphis/S:alfa23 Glu-Gln;beta6Glu-Val. Последствия генных мутаций. Первичный эффект мутации гена проявляется в изменении последовательности аминокислот в полипептидной цепи, кодируемой данным геном. Биологический эффект (на уровне организма) зависит от типа замененной аминокислоты или его расположения в молекуле полипептида. Изменения в структуре или интенсивности синтеза полипептида приводят к нарушениям какого-либо метаболического пути. Таким образом, первичное действие мутантного гена сопровождается многочисленными вторичными эффектами на уровне организма, которые определяют изменения в фенотипе. Мутации могут затрагивать как кодирующую, так и регуляторную часть гена. Изменения в нуклеотидной последовательности в области промотора приводят к количественным изменениям в синтезе мРНК и белка, а именно: - блокированию транскрипции → отсутствию белкового продукта → нарушениям метаболизма (например, в случае фенилкетонурии, галактоземии); - непрерывной активации транскрипции → синтезу белка в избытке → нарушениям в фенотипе (например, избыточный синтез HbFи НЬА2 приводит к гемолизу и анемии). Изменения в нуклеотидной последовательности кодирующей части гена, в особенности, в экзонах, приводят к изменению генетической информации и, соответственно, последовательности аминокислот, вызывая качественные изменения в конечном продукте - белке, например: - синтез белка с пониженной активностью (гипоморфная мутация); - синтез белка с повышенной активностью (гиперморфная мутация); - синтез неактивного белка (аморфная мутация). Исходя из адаптивного значения и влияния их на строение и функции организма, генные мутации можно разделить на: - нейтральные мутации - образуют новые нормальные варианты, увеличивающие внутривидовой биологический полиморфизм (например, группы крови, белки плазмы, тканевые антигены); - отрицательные мутации - приводят к патологическим нарушениям и являются причиной болезней, предрасположенности к болезням; могут нарушать жизнеспособность и воспроизводство, а иногда приводить к смерти носителя из-за несовместимых с жизнью нарушений; - благоприятные мутации - способствуют появлению новых признаков которые позволяют организмам лучше приспособиться к условиям среды.
Множественный аллелизм. В 1902 г. У. Сеттон, а впоследствии Т. Морган сопоставили менделевские законы наследственности с закономерностями поведения хромосом и обнаружили параллелизм между характером наследования генов и распределением хромосом в мейозе. На основании этого они сформулировали хромосомную теорию наследственности. В целом представления школы Т. Х. Моргана можно кратко представить следующим образом: ген имеет основные свойства хромосом (способность к редупликации, к закономерному распределению в митозе и мейозе), занимает определенный участок (локус) хромосомы, является единицей мутации (т. е. изменяется как целое), единицей рекомбинации (т. е. кроссинговера никогда не наблюдали в пределах гена), единицей функции (т. е. все мутации одного гена нарушают одну и ту же функцию). Ген может существовать в двух или нескольких аллельных состояниях. Аллели оказывают различное действие на развитие и фенотипическое выражение признака. Аллелями называют различные состояния одного гена. Как известно, в результате мутирования ген может находиться более чем в двух различных состояниях (явление множественного аллелизма). Поэтому при получении серии мутаций с похожим фенотипом для определения того, затронула мутация один и тот же ген или разные, Морган предложил два теста: функциональный и рекомбинационный. Функциональный критерий основывается на том, что при скрещивании двух мутантов возникает дигетерозигота, имеющая дикий фенотип в силу доминирования нормальных аллелей каждого из генов (мутации комплементарны друг другу). Если скрещиваемые мутанты несут в дигетерозиготе аллельные мутации, то в компаунде дикий тип не появляется, так как оба аллеля одного и того же гена в разных хромосомах имеют мутационные изменения, или, по-другому, мутации не комплементарны. При этом мутации не должны разделяться кроссинговером. (схемка!!!) Например, при скрещивании двух мутантных норок, белой и пастелевой, все гибриды имеют коричневую окраску, т. е. дикий фенотип. При скрещивании белой норки с другой мутантной формой - платиновой - все гибриды имеют платиновую окраску, т. е. мутантный фенотип. Следовательно, в первом случае наблюдается комплементарность, т.е. неаллельность; а во втором — отсутствие комплементарности, т.е. аллельность. В основу рекомбинационного теста было положено представление, что только мутации в разных генах способны рекомбинировать между собой. Исследователи школы Моргана считали мутации аллельными, если соблюдались функциональный (гетерозигота - мутантный фенотип) и рекомбинационный, (рекомбинаций нет) критерии. В связи с изменением представлений о структуре гена уточнялись и критерии аллелизма. Один и тот же ген может изменяться в несколько состояний; иногда таких соетояний бывает несколько десятков и даже сотен. Ген А может мутировать в состояние а1, а2, а3,... аn. Ряд состояний одного и того же гена называют серией множественных аллелей, а само явление — множественным аллелизмом, Изучение серий множественных аллелей показало, что любая аллель такой серии может возникать мутационно непосредственно от аллели дикого типа или любого другого члена данной серии, а каждый из членов серии, по-видимому, имеет свою характерную частоту мутирования. Наследование членов серии множественных аллелей подчиняется менделевским закономерностям. При этом, в отличие от генов, для которых известно только два состояния, сочетание двух разных членов серии множественных аллелей в гетерозиготе называют компаундом. Серии множественных аллелей обнаружены у крупного рогатого скота, кроликов, мышей, морских свинок, дрозофилы, а также у кукурузы, табака, гороха и др. У человека известна серия аллелей: IA, IB, I0 которая определяет полиморфизм по группам крови: Существование серии множественных аллелей локуса, определяющего самостерильность у растений, является тем механизмом, который в ряде случаев обеспечивает перекрестное оплодотворение. Так, было показано, что у табака, клевера и других растений на рыльцах прорастает только пыльца, несущая аллель, отличную от аллелей, имеющихся в генотипе рыльца по локусу самостерильности. Распространенность множественного аллелизма среди животных, растений и микроорганизмов и наличие его у человека могла быть обусловлена тем, что это явление увеличивает резерв мутационной изменчивости, а потому имеет приспособительное значение в эволюции.
26. Спонтанный и индуцированный мутагенез. Мутации, помимо качественных свойств, характеризует и способ возникновения. Спонтанные (случайные) - мутации, возникающие при нормальных условиях жизни. Спонтанный процесс зависит от внешних и внутренних факторов (биологические, химические, физические). Спонтанные мутации возникают у человека в соматических и генеративных тканях. Метод определенияопределения спонтанных мутаций основан на том, что у детей появляется доминантный признак, хотя у его родителей он отсутствует. Проведенное в Дании исследование показали, что примерно одна из 24000 гамет несет в себе доминантную мутацию. Ученый же Холдейн рассчитал среднюю вероятность появления спонтанных мутаций, которая оказалась равна 5*10-5 за поколение. Другой ученый Курт Браун предложил прямой метод оценки таких мутаций, а именно: число мутаций разделить на удвоенное количество обследованных индивидов. Индуцированные мутации Индуцированный мутагенез - это искусственное получение мутаций с помощью мутагенов различной природы. Впервые способность ионизирующих излучений вызывать мутации была обнаружена Г.А. Надсоном и Г.С. Филлиповым. Затем, проводя обширные исследования, была установлена радиобиологическая зависимость мутаций. В 1927 году американским ученым Джозефом Мюллером было доказано, что частота мутаций увеличивается с увеличением дозы воздействия. В конце сороковых годов открыли существование мощных химических мутагенов, которые вызывали серьезные повреждения ДНК человека для целого ряда вирусов. Одним из примеров воздействия мутагенов на человека может служить эндомитоз - удвоение хромосом с последующим делением центромер, но без расхождения хромосом. Возникновение под влиянием ионизирующих излучений и ультрафиолетовых лучей наследственных изменений (мутаций). Под действием излучений возникают качественно те же мутации, что и без облучения, но значительно чаще; соотношение разных типов мутаций также может быть иным. Используется в генетических исследованиях, в селекции промышленных микроорганизмов, сельскохозяйственных и декоративных растений. Повышение частоты вредных мутаций в результате увеличения содержания в биосфере радиоактивных изотопов - одна из основных опасностей радиоактивного загрязнения биосферы. Общая закономерность: с увеличением дозы - увеличивается степень повреждения системы; в процесс вовлекается все большее число составляющих её элементов. На проявление зависимости доза-эффект оказывает существенное влияние внутри- и межвидовая изменчивость организмов. Действительно, особи, относящиеся к одному и тому же виду, существенно отличаются друг от друга по биохимическим, физиологическим, морфологическим характеристикам. Эти отличия в большинстве случаев обусловлены их генетическими особенностями. Еще более выражены, в силу тех же генетических особенностей, межвидовые различия. В этой связи дозы конкретного вещества, в которых оно вызывает повреждение организмов одного и того же и, тем более, разных видов, порой очень существенно различаются. Следовательно, зависимость доза-эффект отражает свойства не только токсиканта, но и организма, на который он действует. На практике это означает, что количественную оценку токсичности, основанную на изучении зависимости доза-эффект, следует проводить в эксперименте на различных биологических объектах, и обязательно прибегать к статистическим методам обработки получаемых данных.
27.Модификационная изменчивость. Пенетрантность и экспрессивность. Норма реакции. Статистические закономерности модификационной изменчивости. Типы модификационных изменений. Рассматривая действие гена, его аллелей, необходимо учитывать не только генные взаимодействия и действие генов-модификаторов, но и модифицирующее действие среды, в которой развивается организм. Известно, что у примулы окраска цветка розовая (Р-) - белая (рр) наследуется по моногибридной схеме, если растения развиваются в интервале температур 15-25°С. Если же растения F2 вырастить при температуре 30-35°С, то все цветки у них оказываются белыми. Наконец, при выращивании растений F2 в условиях температуры, колеблющейся около 30°С, можно получить разнообразные соотношения от 3Р:1рр до 100% растений с белыми цветками. Такое варьирующее соотношение классов при расщеплении в зависимости от условий внешней среды или от условий генотипической среды (так назвал С.С. Четвериков варьирование генотипа по генам-модификаторам) носит название варьирующей пенетрантности: Это понятие подразумевает возможность проявления или непроявления признака у организмов, одинаковых по исследуемым генотипическим факторам. Уже упоминался пример плейотропного действия гена - доминантная платиновая окраска лисиц с рецессивным летальным действием. Как показал Д.К. Беляев с сотрудниками, можно добиться рождения живых щенков, гомозиготных по доминантной аллели платиновой окраски, если варьировать длину дня для беременных самок. Таким образом, пенетрантность проявления летального эффекта может быть снижена (уже не будет 100%-ной). Пенетрантность выражается долей особей, проявляющих исследуемый признак среди всех особей одинакового генотипа по контролируемому (изучаемому) гену. От внешней среды и генов-модификаторов может зависеть и степень выраженности признака. Например, дрозофила, гомозиготная по аллели vgvg (зачаточные крылья), более контрастно проявляет этот признак при понижении температуры. Другой признак дрозофилы - отсутствие глаз (еуеу) варьирует от 0 до 50% от числа фасеток, характерного для мух дикого типа. Степень проявления варьирующего признака называется экспрессивностью. Экспрессивность обычно выражают количественно в зависимости от уклонения признака от дикого типа. Оба понятия - пенетрантность и экспрессивность - были введены в 1925 г. Н.В. Тимофеевым-Ресовским для описания варьирующего проявления генов (рис. 4).
Рис. 4 - Схема, поясняющая варьирование экспрессивности и пенетрантности признака Тот факт, что признак может проявиться или не проявиться у особей данного генотипа в зависимости от условий или варьировать в различных условиях среды, убеждает в том, что фенотип - это результат действия (и взаимодействия) генов в конкретных условиях существования организма. Способность генотипа так или иначе проявляться в различных условиях среды отражает норму его реакции - способность реагировать на варьирующие условия развития. Норму реакции генотипа необходимо учитывать как при экспериментах, так и при выведении новых форм хозяйственно ценных организмов. Отсутствие изменений в проявлении признака указывает на то, что используемое воздействие не влияет на данную норму реакции, а гибель организма - на то, что оно уже за пределами нормы реакции. Селекция высокопродуктивных форм растений, животных и микроорганизмов в значительной степени представляет собой отбор организмов с узкой и специализированной нормой реакции на такие внешние воздействия, как удобрения, обильное кормление, характер выращивания и др. Статистические закономерности модификацнонной изменчивости. Модификационные изменения (модификации) не связаны с изменением генов. Однако модификации могут сильно влиять на их работу, а также на активность ферментов. Хорошо известно, что при низких температурах ферменты гораздо менее активны, что не может не влиять на рост растений и микроорганизмов, развитие животных. Следовательно, действие факторов среды очень существенно для протекания многих физиологических и формообразовательных процессов. Однако эти воздействия, как правило, не влияют на свойства генов, которые передаются в следующие поколения без принципиальных изменений.Именно поэтому модификации не наследуются. Это важное обобщение сделал крупный немецкий биолог А.Вейсман. Примером модификационной изменчивости может служить изменчивость генетически сходных (идентичных) особей. Многие виды растений, например картофель, обычно размножаются вегетативно, в этом случае все потомки обладают одинаковым генотипом. Многие растения существенно отличаются по высоте, кустистости, количеству и форме клубней и другим показателям. Причина этой очень широкой модификационной изменчивости состоит в разнообразном влиянии среды, которое испытывает каждый саженец картофеля. Разнообразие фенотипов, возникающих у организмов под влиянием условий среды, называют модификационной изменчивостью. Спектр модификационной изменчивости определяется нормой реакции.
|
||||
|
|
Последнее изменение этой страницы: 2016-04-23; просмотров: 272; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.41 (0.009 с.) |