
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Кумулятивная и некумулятивная полимерия.Содержание книги
Поиск на нашем сайте
Кумулятивная полимерия. Допустим, что количественные признаки, образующие по своему проявлению непрерывный ряд, определяются взаимодействием многих доминантных генов, действующих на один и тот же признак пли свойство. В таком случае количественно варьирующий признак у разных особей одного и того же поколения будет определяться разным числом доминантных генов в генотипе. Так, при скрещивании рас пшениц с красными и белыми (неокрашенными) зернами шведский генетик Г.Нильсон-Эле в 1908 г. обнаружил в F2 обычное моногибридное расщепление в отношении 3: 1. Однако при скрещивании некоторых других линий пшениц, различающихся по таким же признакам, в F2 наблюдается расщепление в отношении 15/16 окрашенных: 1/16 белых. Окраска зерен из первой группы варьирует от темно- до светло-красной. Интенсивность окраски зерен зависит от числа доминантных генов в генотипе. Гены такого типа, одинаково влияющие на развитие одного признака, были названы генами с однозначным действием,а сами признаки — полимерными.Поскольку эти гены однозначно влияют на один и тот же признак, было принято обозначать их одной латинской буквой с указанием номера разных генов: A1, A2, A3, и т.д. Этот тип взаимодействия генов получил название полимерии. P A1A1A2A2 x a1a1a2a2 красное ↓ белое
F1 A1a1A2a2 красное
Наследование окраски зерна у пшеницы (полимерия) Следовательно, исходные родительские формы, давшие расщепление в F2 15: 1, имели генотипы A1A1A2A2 и a1a1a2a2. Гибрид F1 обладал генотипом A1a1A2a2, а в F2 появились зерна с разным числом доминантных генов. Наличие всех четырех доминантных аллелей A1A1A2A2 у 1/16 растений определяет самую интенсивную окраску зерна; 4/16 всех зерен имели три доминантные аллели (типа A1A1A2a2), 6/16 – две (A1a1A2a2), 4/16 — одну (типа A1a1a2a2). Все эти генотипы определяли различную промежуточную окраску, переходную между интенсивно-красной и белой. Гомозиготной по обоим рецессивным генам (a1a1a2a2) являлась 1/16 всех зерен, и эти зерна оказались неокрашенными. Частоты пяти перечисленных генотипических классов F2 распределяются в ряду: 1 +4 + 6 + 4 + 1 = 16, который отображает изменчивость признака окраски зерна пшеницы в зависимости от числа доминантных аллелей в генотипе. При накоплении таких доминантных генов их действие суммируется, т.е. они имеют кумулятивный эффект, поэтому взаимодействие такого типа называют кумулятивной полимерией.
В опыте Нильсона-Эле тригибридное расщепление в F2 по генам окраски зерен пшеницы давало соотношение 63 красных к 1 неокрашенному. В F2 наблюдались все переходы от интенсивной окраски зерен с генотипом A1A1A2A2A3A3 до полного ее отсутствия у a1a1a2a2a3a3. При этом частоты генотипов с разным количеством доминантных генов распределялись в следующий ряд: 1 + 6 + 15 + 20 + 15 + 6 + 1 = 64. На рисунке приведены гистограммы распределения частот генотипов с разным числом доминантных генов кумулятивного действия в моно-, ди-, три- и полигибридном скрещивании. Из этого сопоставления видно, что, чем большее число доминантных генов определяет данный признак, тем больше амплитуда изменчивости и тем более плавные переходы между различными группами особей. Полимерно наследуется, например, пигментация кожи у человека. При бракосочетании негра и белой женщины рождаются дети с промежуточным цветом кожи (мулаты). У отца и матери мулатов могут родиться дети всех типов кожи с окраской разных оттенков, от черной до белой, что определяется комбинацией двух пар аллелей. Некумулятивная полимерия. Гены с однозначным действием могут определять и качественные, т.е. альтернативные, признаки. Примером может служить наследование оперенности ног у кур. От скрещивания пород, имеющих оперенные и неоперенные ноги, в F1 появляются цыплята с оперенными ногами. Во втором поколении происходит расщепление по фенотипу в отношении 15/16 с оперенными ногами и 1/16 с неоперенными, т.е. наблюдаются два фенотипических класса. Очевидно, порода с оперенными ногами гомозиготна по двум парам доминантных аллелей с однозначным действием (A1A1A2A2), а с неоперенными имеет генотип a1a1a2a2. Гибриды F1 имеют генотип A1a1A2a2. Доминантные аллели каждого из двух генов действуют качественно однозначно, т,е. определяют оперенность ног. Поэтому генотипы A1-A2- (9/16), A1--a2a2- (3/16) и a1a1A2 -(3/16) соответствуют фенотипу с оперенными ногами, а генотип a1a1a2a2 (1/16) – с неоперенными. В приведенном примере наличие в генотипе разного количества доминантных генов однозначного действия не изменяет выраженности признака. Достаточно одной доминантной аллели любого из двух генов, чтобы вызвать развитие признака. Поэтому такой тип взаимодействия генов был назван некумулятивной полимерией. Итак, были разобраны три типа взаимодействия генов: комплементарное, эпистатическое и полимерное. Все они видоизменяют классическую формулу расщепления по фенотипу (9: 3: 3: 1), установленную Менделем для дигибридного скрещивания. Все приведенные типы расщепления по фенотипу столь же закономерны, как 9: 3: 3: 1; они являются не следствием нарушения генетического механизма расщепления, а результатом взаимодействия генов в индивидуальном развитии.
Сцепленное наследование признаков. Генетические доказательства кроссинговера. Цитологические доказательства кроссинговера. Величина кроссинговера и линейное расположение генов в хромосоме. Хромосомная теория наследственности. Современные методы картирования хромосом. Из третьего закона Менделя следует,, что при скрещивании форм, различающихся двумя парами генов (АВ и аb), получается гибрид АaВb, образующий четыре сорта гамет АВ, Аb, аВ и аb в равных количествах. В соответствии с этим в анализирующем скрещивании осуществляется расщепление 1: 1: 1: 1, т.е. сочетания признаков, свойственные родительским формам (АВ и аb), встречаются с такой же частотой, как и новые комбинации (Аb и аВ),— по 25%. Однако по мере накопления фактов генетики все чаще стали сталкиваться с отклонениями от независимого наследования. В отдельных случаях новые комбинации признаков (Аb и аВ) в Fb совсем отсутствовали — наблюдалось полное сцепление между генами исходных форм. Но чаще в потомстве в той или иной степени преобладали родительские сочетания признаков, а новые комбинации встречались с меньшей частотой, чем ожидается при независимом наследовании, т.е. меньше 50%. Таким образом, в данном случае гены чаще наследовались в исходном сочетании (были сцеплены), но иногда это сцепление нарушалось, давая новые комбинации. Совместное наследование генов, ограничивающее их свободное комбинирование, Морган предложил называть сцеплением генов или сцепленным наследованием. Большую работу по изучению наследования неаллельных генов, расположенных в паре гомологичных хромосом, выполнили американский ученый Т.Морган и его ученики. Ученые установили, что гены, расположенные в одной хромосоме, наследуются совместно, или сцеплено. Группы генов, расположенные в одной хромосоме, называют группами сцепления. Сцепленные гены расположены в хромосоме в линейном порядке. Число групп сцепления у генетически хорошо изученных объектов равно числу пар хромосом, то есть гаплоидному числу хромосом. У человека 23 пары хромосом и 23 группы сцепления. Было установлено, что возникают и другие, новыми комбинациями генов, отличающимися от родительской гаметы. Было доказано, что причина возникновения новых гамет заключается в перекресте (кроссинговере) гомологичных хромосом. Гомологичные хромосомы в процессе мейоза перекрещиваются и обмениваются участками. В результате этого возникают качественно новые хромосомы. Частота перекреста между двумя сцепленными генами в одних случаях может быть большой, в других - менее значительной. Это зависит от расстояния между генами в хромосоме. Частота (процент) перекреста между двумя неаллельными генами, расположенными в одной хромосоме, пропорциональна расстоянию между ними. Чем ближе расположены гены в хромосоме, тем теснее сцепление между ними и тем реже они разделяются при перекресте. И наоборот, чем дальше гены отстоят друг от друга, тем слабее сцепление между ними и тем чаще осуществляется перекрест. Следовательно, о расстоянии между генами в хромосоме можно судить по частоте перекреста. Кроссинговер и его генетическое доказательство При допущении размещения в одной хромосоме более одного гена встает вопрос, могут ли аллели одного гена в гомологичной паре хромосом меняться местами, перемещаясь из одной гомологичной хромосомы в другую. Если бы такой процесс не происходил, то гены комбинировались бы только путем случайного расхождения негомологичных хромосом в мейозе, а гены, находящиеся в одной паре гомологичных хромосом, наследовались бы всегда сцепленно — группой. Исследования Т.Моргана и его школы показали, что в гомологичной паре хромосом регулярно происходит обмен генами. Процесс обмена идентичными участками гомологичных хромосом с содержащимися в них генами называют перекрестом хромосом или кроссинговером.Кроссинговер обеспечивает новые сочетания генов, находящихся в гомологичных хромосомах. Явление кроссинговера, так же как и сцепление, оказалось общим для всех животных, растений и микроорганизмов. Наличие обмена идентичными участками между гомологичными хромосомами обеспечивает обмен или рекомбинацию генови тем самым значительно увеличивает роль комбинативной изменчивости в эволюции. О перекресте хромосом можно судить по частоте возникновения организмов с новым сочетанием признаков. Такие организмы называют рекомбинантами. Рассмотрим один из классических опытов Моргана на дрозофиле, позволивший ему доказать, что гены расположены в хромосомах в определенном порядке. Когда гены находятся в разных парах хромосом, то, например, генотип дигетерозиготы записывается так: У дрозофилы рецессивный ген черной окраски тела обозначается b, а его доминантная аллель, определяющая дикую серую окраску, -b+, ген рудиментарных крыльев - vg, нормальных – vg+. При скрещивании мух, различающихся по двум парам сцепленных признаков, серых с рудиментарными крыльями Если гибридные самцы скрещиваются с самками, гомозиготными по обоим рецессивным генам (♀ Гаметы с хромосомами, претерпевшими кроссинговер, называют кроссоверными,а с непретерпевшими — некроссоверными.Соответственно организмы, возникшие от сочетания кроссоверных гамет гибрида с гаметами анализатора, называют кроссоверамиили рекомбинантами,а возникшие за счет некроссоворных гамет гибрида — некроссоверными или нерекомбинантными. Цитологическое доказательство кроссинговера После того как генетическими методами удалось установить явление кроссинговера, необходимо было получить прямое доказательство обмена участками гомологичных хромосом, сопровождающегося рекомбинацией генов. Наблюдаемые в профазе мейоза картины хиазм могут служить лишь косвенным доказательством этого явления, констатация происшедшего обмена прямым наблюдением невозможна, так как обменивающиеся участками гомологичные хромосомы обычно абсолютна одинаковы но величине и форме. Чтобы сопоставить цитологические карты гигантских хромосом с генетическими, Бриджес предложил воспользоваться коэффициентом кроссинговера.Для этого он разделил общую длину всех хромосом слюнных желез (1180 мкм) на общую длину генетических карт (279 единиц). В среднем это отношение оказалось равным 4,2. Следовательно, каждой единице перекреста на генетической карте соответствует 4,2 мкм на цитологической карте (для хромосом слюнных желез). Зная расстояние между генами на генетической карте какой-либо хромосомы, можно сравнить относительную частоту перекреста в разных ее районах. Например, в Х- хромосоме дрозофилы гены у и ec находятся на расстоянии 5,5%, следовательно, расстояние между ними в гигантской хромосоме должно быть 4,2 мкм Х 5,5 = 23 мкм, но непосредственное измерение дает 30 мкм. Значит, в этом районе Х -хромосомы кроссинговер идет реже средней нормы. В силу неравномерного осуществления обменов по длине хромосом гены при нанесении их на карту распределяются на ней с разной плотностью. Следовательно, распределение генов на генетических картах можно рассматривать как показатель возможности осуществления перекреста по длине хромосомы. При анализе расщепления в случае кроссинговера обращает на себя внимание определенное количественное отношение кроссоверных и некроссоверных классов. Обе исходные родительские комбинации признаков, образовавшиеся из некроссоверных гамет, оказываются в потомстве анализирующего скрещивания в равном количественном отношении. В указанном опыте с дрозофилой тех и других особей было примерно по 41,5%. В сумме некроссоверные мухи составили 83% от общего числа потомков. Два кроссоверных класса по числу особей также одинаковы, и сумма их равна 17%. Частота кроссинговера не зависит от аллельного состояния генов, участвующих в скрещивании. Если в качестве родителя использовать мух Результаты этих опытов показывают, что сцепление генов реально существует, и лишь в известном проценте случаев оно нарушается вследствие кроссинговера. Отсюда и был сделан вывод, что между гомологичными хромосомами может осуществляться взаимный обмен идентичными участками, в результате чего гены, находящиеся в этих участках парных хромосом, перемещаются из одной гомологичной хромосомы в другую. Отсутствие перекреста (полное сцепление) между генами представляет исключение и известно лишь у гетерогаметного пола немногих видов, например у дрозофилы и шелкопряда. Изученное Морганом сцепленное наследование признаков получило название закона сцепления Моргана.Поскольку рекомбинация осуществляется между генами, а сам ген кроссинговером не разделяется, его стали считать единицей кроссинговера. Положение хромосомной теории: Каждый ген имеет своё строго определённое положение в хромосоме. Гены расположены в хромосоме линейно в строго определённом порядке. Причиной появления особей с перекомбенированными признаками является кроссенговер. Чем дальше гены друг от друга расположены в хромосоме, тем больше вероятность кроссенговера между ними.
Т. Морган предположил, что гены расположены в хромосомах линейно, а частота кроссинговера отражает относительное расстояние между ними: чем чаще осуществляется кроссинговер, тем далее отстоят гены друг от друга в хромосоме; чем реже кроссинговер, тем они ближе друг к другу. Одним из классических опытов Моргана на дрозофиле, доказывающим линейное расположение генов, был следующий. Самки, гетерозиготные по трем сцепленным рецессивным генам, определяющим желтую окраску тела y, белый цвет глаз w и вильчатые крылья bi, были скрещены с самцами, гомозиготными по этим трем генам. В потомстве было получено 1,2% мух кроссоверных, возникших от перекреста между генами у и w; 3,5% − от кроссинговера между генами w и bi и 4,7% — между у и bi. Из этих данных с очевидностью вытекает, что процент перекреста является функцией расстояния между генами. Поскольку расстояние между крайними генами у и bi равно сумме двух расстояний между у и w, w и bi, следует предположить, что гены расположены в хромосоме последовательно, т.е. линейно. Воспроизводимость этих результатов в повторных опытах указывает на то, что местоположение генов в хромосоме строго фиксировано, т. е. каждый ген занимает в хромосоме свое определенное место — локус. Основным положениям хромосомной теории наследственности — парности аллелей, их редукции в мейозе и линейному расположению генов в хромосоме — соответствует однонитчатая модель хромосомы Приняв положения, что генов в хромосоме может быть много и расположены они в хромосоме в линейном порядке, а каждый ген занимает определённый локус в хромосоме, Морган допустил, что перекрест между гомологичными хромосомами может происходить одновременно в нескольких точках. Это предположение было им доказано тоже на дрозофиле, а затем полностью подтвердилось на ряде других животных, а также на растениях и микроорганизмах. Кроссинговер, происходящий лишь в одном месте, называют одинарным, в двух точках одновременно – двойным, в трёх – тройным и т.д., т.е. он может быть множественным. Чем дальше отстоят друг от друга в хромосоме гены, тем больше вероятность двойных перекрестов между ними. Процент рекомбинаций между двумя генами тем точнее отражает расстояние между ними, чем оно меньше, так как в случае малого расстояния уменьшается возможность двойных обменов. Для учета двойного кроссинговера необходимо иметь дополнительный маркер, находящийся между двумя изучаемыми генами. Определение расстояния между генами осуществляют следующим образом: к сумме процентов одинарных кроссоверных классов прибавляют удвоенный процент двойных кроссинговеров. Удвоение процента двойных кроссинговеров необходимо в связи с тем, что каждый двойной кроссинговер возникает благодаря двум независимым одинарным разрывам в двух точках.
Установлено, что кроссинговер, происшедший в одном месте хромосомы, подавляет кроссинговер в близлежащих районах. Это явление носит название интерференции.При двойном перекресте интерференция проявляется особенно сильно в случае малых расстояний между генами. Разрывы хромосом оказываются зависимыми друг от друга. Степень этой зависимости определяется расстоянием между происходящими разрывами: по мере удаления от места разрыва возможность другого разрыва увеличивается. Эффект интерференции измеряется отношением числа наблюдаемых двойных разрывов к числу возможных при допущении полной независимости каждого из разрывов. Объясним это на рассмотренном ранее примере с кукурузой. На основании измерения частоты перекреста было установлено, что в генотипе
Но в опыте получено всего 1,5% растений, возникших вследствие двойного кроссинговера, т.е. ниже ожидаемого, что и объясняется наличием интерференции. Величину интерференции измеряют отношением наблюдаемого числа двойных перекрестов к теоретически ожидаемому. Это отношение в генетике называют коинциденциейи выражают в долях единицы или в процентах. В приведенном примере она равна 1,5: 2,5, т.е. 0,6, или 60%. На небольшом расстоянии, где имеет место влияние одного разрыва на другой, коинциденция будет меньше 1. Но влияние интерференции распространяется лишь на определенное расстояние, а затем исчезает. В последнем случае коинциденция равна 1, или 100%. Коинциденция в разных хромосомах и в разных участках одной и той же хромосомы различна. Особенности распределения генов в хромосомах, их строение, а также расположение центромеры влияют на частоту перекреста хромосом. Подробно об этом будет сказано дальше.
|
|||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2016-04-23; просмотров: 765; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.137 (0.014 с.) |

 Если у гибридов F1 таких генов в гетерозиготном состоянии оказывается не два, а три (A1a1A2a2A3a3) или более, то число комбинаций генотипов в F2 увеличивается. Этот ряд генотипов можно представить в виде биномиальной кривой изменчивости данного признака.
Если у гибридов F1 таких генов в гетерозиготном состоянии оказывается не два, а три (A1a1A2a2A3a3) или более, то число комбинаций генотипов в F2 увеличивается. Этот ряд генотипов можно представить в виде биномиальной кривой изменчивости данного признака. . Если гены находятсяв одной паре гомологичных хромосом, формула видоизменяется:
. Если гены находятсяв одной паре гомологичных хромосом, формула видоизменяется:  . При этом аллели одного гена (Аа и Bb), находящиеся в гомологичных хромосомах, пишутся строго одна под другой.
. При этом аллели одного гена (Аа и Bb), находящиеся в гомологичных хромосомах, пишутся строго одна под другой. и черных с нормальными крыльями
и черных с нормальными крыльями  − гибриды F1
− гибриды F1  серые с нормальными крыльями.
серые с нормальными крыльями. ♂
♂  ),
),  то в потомстве получается расщепление в отношении 1 серотелая муха с рудиментарными крыльями: 1 чернотелая с нормальными крыльями. Следовательно, данная дигетерозигота образует только два сорта гамет (b+vg и bvg+) вместо четырех, причем сочетание генов в гаметах самца соответствует тому, которое было у его родителей. Исходя из указанного расщепления, следует предположить, что у самца не происходит обмен участками гомологичных хромосом. Действительно, у самцов дрозофилы как в аутосомах, так и в половых хромосомах, кроссинговер в норме не происходит, благодаря чему наблюдается полное сцепление генов, находящихся в одной хромосоме.
то в потомстве получается расщепление в отношении 1 серотелая муха с рудиментарными крыльями: 1 чернотелая с нормальными крыльями. Следовательно, данная дигетерозигота образует только два сорта гамет (b+vg и bvg+) вместо четырех, причем сочетание генов в гаметах самца соответствует тому, которое было у его родителей. Исходя из указанного расщепления, следует предположить, что у самца не происходит обмен участками гомологичных хромосом. Действительно, у самцов дрозофилы как в аутосомах, так и в половых хромосомах, кроссинговер в норме не происходит, благодаря чему наблюдается полное сцепление генов, находящихся в одной хромосоме. и
и 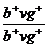 , то в анализирующем скрещивании кроссоверные (b+vg и bvg+) и некроссоверные (bvg и b+vg+) особи появятся с той же частотой (соответственно 17 и 83%), что и в первом случае.
, то в анализирующем скрещивании кроссоверные (b+vg и bvg+) и некроссоверные (bvg и b+vg+) особи появятся с той же частотой (соответственно 17 и 83%), что и в первом случае. гены v и gl разделяются расстоянием 18,3%, а gl и sl — 13,6%. Если разрывы на участках v — gl и gl — sl происходят как независимые друг от друга события, то вероятность двойного кроссинговера между генами v и sl должна быть равна произведению процентов кроссинговера на участках v — gl и gl — sl, т.е.
гены v и gl разделяются расстоянием 18,3%, а gl и sl — 13,6%. Если разрывы на участках v — gl и gl — sl происходят как независимые друг от друга события, то вероятность двойного кроссинговера между генами v и sl должна быть равна произведению процентов кроссинговера на участках v — gl и gl — sl, т.е.



