Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Накопление и выделение энергииСодержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте В живых организмах энергия накапливается и высвобождается множеством разных способов, одни из которых более широко распространены, другие – менее. Рассмотрим вначале, каким образом выделяется энергия, запасённая в клетке. Там энергия аккумулируется в форме сахаров, полисахаридов, некоторых липидов, а иногда и белков. Она получается из молекул этих веществ при их превращении в меньшие молекулы (катаболизм). Главным поставщиком клеточной энергии служит АТФ (разд. 2.2.2). При переходе молекулы АТФ в родственную ей, но более простую молекулу АДФ высвобождается энергия. И наоборот, при поглощении энергии АДФ переходит в АТФ. Заметим, что АТФ – лишь временный хранитель энергии и действует как молекулярное связующее звено между энергией, освобождающейся при распаде молекул в накопителе, и энергией, требующейся для протекания некоторого процесса. Для превращения одной молекулы АДФ в АТФ необходима энергия 5,1 ´ 10–20 Дж (джоуль) – это удобная единица измерения энергии клетки, которая может также выражаться числом образующихся молекул АТФ. Почти у всех организмов катаболизм проявляется в виде процесса дыхания. Для аэробного дыхания нужен кислород – вторая по распространённости составляющая земной атмосферы, присутствующая также в воде рек, озёр и океанов. Большинство эукариотов используют кислород в химической реакции по превращению органических веществ данного носителя энергии в воду и диоксид углерода; в ходе этой реакции высвобождается энергия, идущая на образование АТФ из АДФ. Если носителем является глюкоза, то длинный ряд химических реакций можно в суммарном виде представить как C6H12O6 + 6O2 ® 6CO2 + 6H2O (2.4.) Это пример окислительно-восстановительной реакции, в которой топливом служит глюкоза, а окислителем – кислород. У организмов, живущих в отсутствии кислорода, важную роль играет другой тип дыхания – так называемое анаэробное дыхание, которое в наши дни характерно в той или иной его форме для самых разных одноклеточных организмов. Примером такого процесса служит брожение – образование спирта под действием дрожжей, представляющих собой колонии одноклеточных эукариотов, принадлежащих к царству грибов. Брожение приводит к образованию АТФ из АДФ благодаря энергии, выделяющейся в процессе превращения глюкозы в этанол (этиловый спирт) и диоксид углерода. Конечный результат нескольких стадий выглядит следующим образом: С6Н12О6 ® 2СО2 + 2С2Н5ОН (2.5) Однако из одной молекулы глюкозы образуется только две молекулы АТФ, тогда как при аэробном дыхании возникает 36 молекул АТФ.
Рис 2.6 Организмы, в жизнедеятельности которых принимает участие кислород, называются аэробами. Остальные относятся к анаэробам: для некоторых из них кислород губителен, другие же в присутствии кислорода используют аэробное дыхание. На рис. 2.6 представлены эти два вида дыхания. Там же сведены воедино различные пути накопления клеточной энергии, к знакомству с которыми мы сейчас и перейдём.
Накопление энергии
Откуда берутся сахара, полисахариды, липиды и, иногда, белки, благодаря которым в клетке создаются запасы энергии? Для некоторых одноклеточных и множества многоклеточных организмов, включая животных, ответом является «пища» (т.е. органические вещества других живых существ, которые они заглатывают или впитывают, хотя и возможна их некоторая предварительная обработка). Особенно всеядны люди, получающие органическое вещество из множества самых разных источников. Организмы, чья жизнедеятельность основана на таком уже готовом органическом материале, называют гетеротрофами («поедателями других»). Если же организмы синтезируют эти вещества из неорганических соединений, их называют автотрофами («кормящий себя»); они располагаются в основании пищевой цепи. К автотрофам относятся зелёные растения, вырабатывающие органическое вещество в процессе фотосинтеза, при котором из СО2 и воды образуется глюкоза. СО2 служит источником углерода. Хотя этот процесс достаточно сложный и состоит из нескольких стадий, в конечном счёте в зелёных растениях получается следующий результат: 6СО2 + 6Н2О ® С6Н12О6 + 6О2, (2.6) где СО2 восстанавливается в форме глюкозы, а электроны берутся у Н2О, окисление которой даёт кислород. Поскольку побочным продуктом здесь является кислород, этот процесс называют кислородным фотосинтезом. Продукты (в правой части уравнения реакции) обладают большей энергией, чем реагенты (в левой части), и поэтому для протекания реакции необходим источник энергии. В его роли выступает солнечный свет. Солнечное излучение, как и любое другое электромагнитное излучение, распространяется в космическом пространстве в виде волн с частотой n и длиной волны l, которые связаны между собой соотношением с = ln, где с – скорость волны (в данном случае это скорость света). Но когда электромагнитное излучение вступает во взаимодействие с веществом, оно скорее напоминает поток частиц, называемых фотонами. Энергия E фотона пропорциональна частоте волны, и существует соотношение E = hn = hc/l,(2.7) где h – универсальная константа, называемая постоянной Планка (см. учебники по физике, указанные в Приложениях). В зелёных растениях молекула хлорофилла поглощает два фотона в интервале длин волн 0,4 – 0,7 мкм, и это приводит к целому ряду явлений. Важным промежуточным этапом среди них является превращение АДФ в АТФ и образование вещества под названием НАДФ·Н2 (детали этого процесса для нас не играют роли). Из этих двух веществ затем получается глюкоза, и таким образом создаётся запас энергии. Образуются также и другие вещества, например аминокислоты. У некоторых одноклеточных организмов тоже происходит фотосинтез примерно таким же образом. У ряда прокариотов это происходит иначе. Они осуществляют фотосинтез, используя в качестве донора электронов не Н2О, а такие молекулы, как сероводород (Н2S), и кислород при этом не образуется. Схематически этот процесс можно представить в следующем виде: 2H2S + CO2 + (C) ® 2 "CH2O" + 2S, (2.8) где СН2О – простейший углевод, показанный в кавычках, поскольку образуемые в реальности углеводы сложнее. Это менее эффективный процесс, чем кислородный фотосинтез, так как он потребляет меньшее количество энергии поступающего солнечного излучения, хотя и может использоваться организмами, нетерпимыми к кислороду. И всё-таки процессы биосинтеза и дыхания сильно выиграли от появления кислорода в земной атмосфере, поскольку благодаря ему появилась возможность использовать больше энергии. Некоторые автотрофные прокариоты накапливают энергию в результате химических реакций без участия фотосинтеза. Этот процесс называется хемосинтезом. При хемосинтезе организм использует вещества из окружающей среды, которые могут вступать в экзотермические реакции внутри клетки. Выделяемая энергия идёт на образование АТФ и т.д. На глубине более 100 м даже в чистой воде из-за недостатка солнечного света фотосинтез уже не может происходить, а возможен только хемосинтез. То же самое можно сказать и о подземных полостях и трещинах. Особенно важную роль, благодаря их энергетическому выходу, играют окислительно-восстановительные реакции. Одним из многочисленных примеров может служить пара растворённых газов СО2 и Н2, выделяющихся из жерл вулканов на океаническом дне. При температурах около 400 °С эти газы остаются нетронутыми, но при более низких температурах они вступают друг с другом в реакцию следующим образом: СО2 + 4Н2 ® СН4 + 2Н2О. (2.9) Однако скорость этой реакции крайне мала, и поэтому они продолжают существовать, и некоторые клетки имеют шанс их поглотить. Внутри клетки скорость реакции существенно возрастает под действием ферментов (разд. 2.2.1). Суммарный эффект состоит в образовании углевода ‘CH2O’ как носителя энергии, причём источником углерода снова служит СО2: 2СО2 + 6Н2 ® ‘CH2O’ + CH4 + 3H2O. (2.10) Из-за появления СН4 в ходе этого процесса последний приобрел название метаногенез, а осуществляющие его организмы называют метаногенами. На рис. 2.6 изображены эти разнообразные пути накопления энергии в живых клетках. Рассмотрим теперь основные процессы, для которых требуется эта энергия.
2.4.3. Синтез белка
Если не считать воду, а также клеточные стенки у некоторых организмов, то в наибольшей степени масса клетки создаётся белками (см. табл. 2.2). Кроме того, они выполняют наибольшее количество функций, но молекулы лишь очень немногих белков могут сами себя воспроизводить. Поэтому, чтобы образовался белок, почти неизменно требуется что-то ещё, а именно ДНК, которая находится в клетке. ДНК содержит программы по созданию всех клеточных белков и потому является носителем всей генетической информации. Синтез белков в большинстве клеток потребляет больше энергии, чем любой другой процесс биологического синтеза.
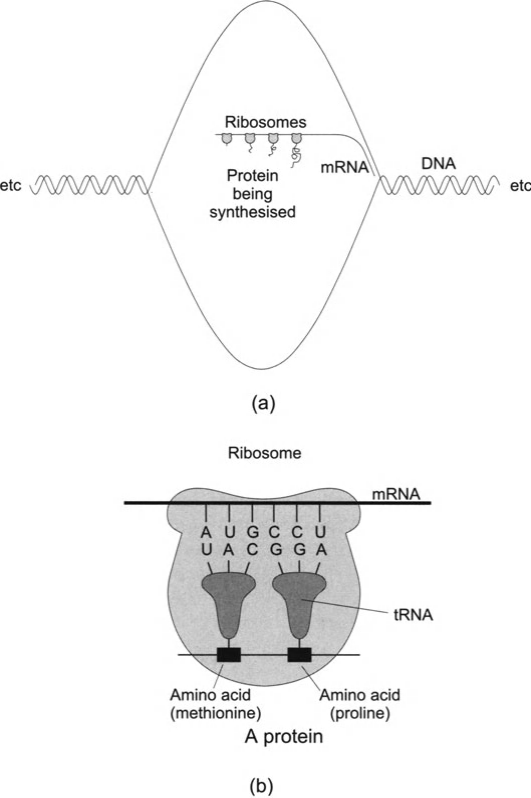
Рис 2.7 Рассмотрим типичную прокариотическую клетку. Синтез белка происходит в два основных этапа. Во-первых, часть молекулы ДНК разделяется под действием фермента, который разрывает слабые связи между парами оснований, образующих ступеньки молекулярной лестницы. Определённый фермент может разорвать связи от определённого места старта до определённого места остановки. Последовательность оснований между этими крайними точками составляет ген. Только на одной из двух нитей разрушенной ДНК фермент строит РНК, как показано на рис. 2.7(а); при этом используются основания РНК, свободно плавающие в клетке. Заметим, что в результате образуется определённая форма РНК, в которой последовательность оснований является зеркальным отражением последовательности на нити ДНК, согласно правилам, по которым А(ДНК) образует пару с U(РНК), С(ДНК) – c G(РНК), G(ДНК) – с С(РНК) и T(ДНК) – с А(РНК) (рис. 2.7(б)). Когда формирование этой РНК завершено, фермент и РНК освобождают ДНК, которая восстанавливается. Образуемая таким путём РНК называется матричной РНК (мРНК). Второй этап – формирование определённой цепи аминокислот, т.е. конкретного белка, соответствующего данной последовательности оснований мРНК. Этот процесс происходит внутри рибосом (рис. 2.7(а) и (б)). Основания (по три в группе) располагаются вдоль мРНК, определяя последовательность аминокислот в данном белке и обусловливая тем самым тип белка. Каждый триплет оснований называется кодоном и соответствует только одной из приблизительно 20 различных аминокислот. Форма РНК, называемая транспортной РНК (тРНК), является связующим звеном между кодоном мРНК и аминокислотой. Для каждой аминокислоты существует один вид тРНК. В процессе синтеза белка в рибосоме одномоментно содержится только часть молекулы мРНК и до двух молекул тРНК, которые рибосома на время захватывает из цитоплазмы с целью подобрать их в соответствии с тем участком мРНК, который в тот момент находится внутри неё. Процесс начинается с присоединения молекулы тРНК, несущей на себе аминокислоту, к молекуле мРНК в соответствии с первым кодоном. Затем другая молекула тРНК присоединяется к мРНК на следующем кодоне – это тРНК иного типа, если только кодон не является таким же, как в первом случае. Так соответствующая аминокислота присоединяется к первой. После этого первая молекула тРНК отделяется, присоединяется третья и т.д. до тех пор, пока не синтезируется весь белок. Рибосомы взаимодействуют с мРНК, пока не закончится образование белка. Весь процесс повторяется для других генов вплоть до синтеза всех необходимых белков. Сама рибосома состоит из белков и молекул третьей формы РНК, называемых рибосомными РНК (рРНК). Возможно, именно рРНК служит катализатором всей сборки, а не рибосомные белки. В эукариотических клетках основные биохимические процессы синтеза белка с участием ДНК и РНК в целом те же, что и в прокариотических, хотя и несколько сложнее. У гетеротрофных организмов – как прокариотов, так и эукариотов, большинство аминокислот поступает из пищи и проходит в клетку через мембрану, хотя животные могут синтезировать некоторые из них (в митохондриях – см. разд. 2.3). У автотрофов они синтезируются внутри клетки.
2.4.4. Воспроизведение и эволюция
Все живые организмы имеют ограниченную продолжительность жизни. Поэтому, чтобы сохраниться, каждый конкретный вид должен обладать способностью к самовоспроизведению. У всех современных организмов информация, необходимая для синтеза всех его разнообразных белков, полностью содержится в молекуле ДНК. Поэтому информация для создания копии его самого также находится в молекуле ДНК. В связи с этим не приходится удивляться, что для продолжения существования организма от одного поколения к другому требуется удвоение ДНК.
Рис 2.8 На рис. 2.8 в сильно упрощённом виде показаны основные стадии деления прокариотической клетки. На первой стадии ДНК самовоспроизводится способом, который мы ещё вкратце опишем. После этого клетка имеет уже две одинаковые молекулы ДНК, которые перемещаются к противоположным полюсам клетки. Вторая стадия наступает, когда у клетки развивается «перетяжка» из новой мембраны, которая делит клетку пополам. На последней стадии такая клетка разделяется на две (более или менее) идентичные клетки. Каждая из этих новых клеток может подвергаться дальнейшему удвоению.
Рис 2.9 Вопрос о том, что вызывает репликацию ДНК, мы оставим за пределом этой книги. Результат же состоит в том, что определённые ферменты постепенно разделяют молекулу ДНК на две её нити (рис. 2.9). Получаются две группы открытых непарных оснований. Каждое открытое основание притягивает свои комплементарные нуклеотиды из числа свободных нуклеотидов, в изобилии присутствующих в клетке. Эти свободные нуклеотиды будут заново синтезированы благодаря действию ферментов. Чтобы связать их для формирования новой ДНК, нужен ещё один фермент. Таким образом, из каждой нити первоначальной молекулы ДНК строится одна полная молекула ДНК. Последовательность оснований в обеих молекулах одна и та же. Весь процесс репликации ДНК осуществляется ферментами. Таким образом, ферменты участвуют в построении ДНК, но и сами, будучи белками, нуждаются в ДНК для своего синтезирования. Когда придёт время рассмотреть происхождение жизни (в гл. 3), нам нужно будет обратить внимание на эту "проблему курицы и яйца". При делении клетки репликация ДНК не всегда протекает строго идеально. Поэтому «дети» не идентичны своим «родителям». Если «дети» оказываются жизнеспособными, разнообразие только возрастает. Оно может возникать также из-за переноса части ДНК прокариотической клетки в другую прокариотическую клетку, возможно, совсем иного типа, с включением этой ДНК в ДНК реципиента. Такой перенос осуществляется разными способами. Другой источник изменений – повреждение ДНК под действием каких-то химических соединений, имеющихся в окружающей среде, а также солнечного ультрафиолета, заряженных частиц высоких энергий и гамма-излучения радиоактивных веществ, находящихся на Земле и в космосе. Ещё одной причиной служат внутренние клеточные процессы, заставляющие некоторые участки ДНК перемещаться с одного места на другое. Все эти изменения ДНК мы называем мутациями. Деление происходит и в эукариотических клетках, хотя соответствующие процессы здесь отличаются от случая прокариотов. Один процесс (называемый митозом) обеспечивает размножение одноклеточных эукариотов неполовым путём и позволяет многоклеточным организмам расти и замещать вышедшие из строя или утраченные клетки. Однако репродукция организма может происходить и по-другому. У растений и животных она происходит путём полового размножения, при котором половина ДНК для нового организма берётся из специфических клеток, имеющихся в особи женского пола, а другая половина – из специфических клеток мужской особи (у животных это соответственно яйцо и сперматозоид). Отсюда следует, что ДНК потомков не совпадает с ДНК обоих родителей, хотя и является весьма похожей, что помогает нам отнести потомка к тому же виду организмов, что и его родителей. Деление эукариотической клетки, после которого происходит объединение ДНК двух родителей, называется мейозом. Половое размножение явно стимулирует генетические изменения в ДНК потомства. Дальнейшие вариации у эукариотов возникают вследствие мутаций молекулы ДНК. Таким образом, в организмах любого типа – и у прокариотов, и у эукариотов – ДНК не является чем-то неизменным, переходящим от одного поколения к другому. В подавляющем большинстве случаев изменения ДНК не приводят ни к какому-либо заметному эффекту, ни к важным последствиям или каким-то повреждениям. Но одни потомки оказываются лучше приспособленными к окружающей среде, чем другие, и именно они получают больший шанс на выживание и самовоспроизведение. Это и сделало возможной эволюцию жизни от её самых ранних форм, и направило её по тому пути, который приводит к выживанию вида и появлению новых жизнеспособных видов. Эволюция происходит благодаря разнообразию потомства. Поэтому наследуемые свойства, способствующие выживанию и воспроизведению, передаются от поколения к поколению. Такова основа теории Дарвина об эволюции путём естественного отбора, и она ведёт к разнообразию видов, каждый из которых приспособлен к условиям окружающей его среды.
2.5. Многообразие сред обитания
Живые организмы встречаются на Земле повсеместно – на поверхности суши и в океанах. Они присутствуют также в земной коре – не только в подземных полостях и осадках, но и внутри самих горных пород. Таким образом, живые существа населяют самые различные среды, хотя в наиболее экстремальных условиях существуют главным образом прокариоты. Но, несмотря на такое многообразие условий, во всех средах обитания обычно присутствует жидкая вода. Чтобы выжить, организм должен иметь доступ к воде и существовать в тех условиях, при которых эта вода внутри его клеток может находиться в жидком состоянии. Никакой организм не может выжить, если в течение хотя бы части его жизненного цикла он не имеет в своём составе воды в жидкой фазе. Подавляющее большинство живых организмов нуждается в получении воды извне, причем в жидком состоянии, а не в виде льда или пара. Рассмотрим вначале обычные (неэкстремальные) среды обитания.
2.5.1. Обычные условия обитания
Самым знакомым для нас местом обитания, конечно, является земная поверхность. Она соприкасается с богатой кислородом земной атмосферой (см. табл. 2.1), где среднее давление на уровне моря равно 1,013 ´ 105 Па, а глобальная средняя температура поверхности 15 °С (в большинстве районов Земли в течение большей части времени она заключена между 0 и 35 °С). Если бы у человека не было такой защиты, как одежда и жилище, он мог бы выжить лишь в очень узком диапазоне условий окружающей среды. Даже при наличии обильных запасов пищи и жидкой воды мы умерли бы, если бы нам пришлось всего несколько дней выносить температуру вне интервала примерно от 5 до 45 °С. Мало животных и растений могут жить при длительном воздействии температур за пределами диапазона 0 – 45 °С, а при температурах примерно ниже –20 и выше 50 °С – вообще никто. Для выживания некоторых организмов имеет значение также величина парциального давления О2 в местной атмосфере. Для человека нижним пределом выживания является парциальное давление кислорода около 0,04 ´ 105 Па, т.е. примерно в пять раз меньшее, чем на уровне моря, и встречается оно на высоте около 12 км. Однако многие места обитания вообще лишены кислорода, скажем озёрные осадки, где существуют анаэробные организмы (разд.2.4.2). В самом деле, на ранней стадии существования жизни на Земле, как мы увидим в разд. 3.2.3, в атмосфере было так мало кислорода, что аэробы не могли жить в таких условиях, и выживали только анаэробы. Поэтому мы не станем относить отсутствие кислорода к экстремальным условиям окружающей среды. К числу ограничивающих факторов относятся также солёность и щёлочность–кислотность окружающей среды. Мерой щёлочности–кислотности служит величина рН. Нейтральная среда типа чистой воды характеризуется рН = 7,0, щелочные среды имеют рН > 7,0, а кислые рН < 7,0. Таким образом, обычно используемый в быту раствор стиральной соды имеет рН» 10,5 и является слабой щёлочью, а купленный в гастрономе уксус характеризуется рН = 2 ¸ 3, т.е. это слабая кислота. В подавляющем большинстве случаев внутри клетки рН = 7,7, а значит, среда является мягкой щёлочью и должна сохраняться такой независимо от величины рН окружающей среды. Точно так же солёность внутри клетки должна составлять примерно одну треть от солёности морской воды. Как правило, солёность определяется содержанием NaCl; в меньших, но измеримых количествах там присутствуют и другие соли. В клетке относительное содержание NaCl по массе около 0,85%. Помимо этого для всех форм жизни облучение ультрафиолетом и другими видами ионизирующего излучения не должно превышать некоторые пределы: в противном случае могут возникнуть слишком сильные повреждения ДНК и других молекул.
2.5.2. Экстремальные условия обитания
В табл. 2.3 приведены те экстремальные значения характеристик окружающей среды, при которых обитают различные земные организмы; в некоторых случаях при ещё более широком диапазоне параметров выживание происходит в состоянии, близком к анабиозу. Вряд ли найдется организм, способный выжить во всём объединённом диапазоне условий. Каждый из диапазонов, как правило, относится только к определённым организмам. При большинстве крайних значений параметров обнаружены лишь одноклеточные организмы, причём наибольшая их часть – прокариоты. Они гораздо лучше переносят экстремальные условия окружающей среды, чем растения и животные. Отчасти это объясняется тем, что последние состоят из клеток многих типов, имеющих специализированные функции, а также из различных органов и крупномасштабных структур. Поддерживать всё это в рабочем состоянии обычно удаётся только при умеренных условиях. Организмы, живущие в экстремальных условиях, называют экстремофилами («любителями экстрима»), хотя это выражение отдаёт шовинизмом: получается, что именно мы сами живём в оптимальных условиях.
Таблица 2.3. Экстремальные условия среды обитания земных организмов (главным образом прокариотов).
Поразительны крайние значения диапазонов, приведенные в табл. 2.3. При замерзании и кипении воды клетка разрушается – тогда как же некоторые организмы могут выживать при –18 °С, а другие – при 123 °С? Но знакомые нам значения температур замерзания и кипения, 0 и 100 °С, относятся только к чистой воде при давлении 1,013 ´ 105 Па.
Рис 2.10 На рис. 2.10 приведены интервалы значений температуры и давления, при которых чистая вода может долго существовать в жидком, твёрдом и газообразном состояниях. Если давление превышает 1,013 ´ 105 Па, точка кипения повышается. Этот случай как раз соответствует гипертермофилам («любителям экстремальной жары»), обнаруженным при высоких давлениях в глубинах океанов. В условиях достаточно высокого давления верхний предел температур, при которых может существовать жизнь, основанная на углероде и жидкой воде, в принципе может достигать 160 °С, т.е. значения, при которых важнейшие соединения углерода не могут не разрушаться. При другом крайнем значении температуры психрофилы («любители холода») должны избегать трёх опасностей. Ясно, что вода в клетке не должна замерзать. Давление тут не поможет – как видно из рис. 2.10, оно практически не влияет на точку замерзания. Но если в воде растворены какие-либо вещества, то температура её замерзания может существенно понизиться (а температура кипения опять-таки повысится). В клетках психрофилов присутствие некоторых белков понижает точку замерзания воды. Далее, скорости биохимических реакций не должны становиться слишком низкими. При падении температуры они экспоненциально уменьшаются, и в других организмах – не-психрофилах – становятся крайне малы, когда температура опускается ниже приблизительно 10 °С. Психрофилы обходят эту проблему с помощью специальных ферментов, которые не обнаружены в других клетках; они способствуют протеканию реакций при низких температурах. Наконец, клеточная мембрана, чтобы регулировать потоки проходящих через неё веществ, не должна становиться слишком вязкой. Это решается включением в мембрану особых липидов, благодаря которым она остаётся гибкой. Нижнее предельное значение давления в табл. 2.3 следует из того, что вода должна быть жидкой. Для чистой воды минимальное давление равно 610 Па; при более низких его значениях вода может пребывать только в твёрдом или газообразном состояниях (рис. 2.10). Верхний конец интервала давлений – гидростатическое давление в самом глубоком месте океана, где обнаружена жизнь (Марианский жёлоб в западной части Тихого океана). Для одноклеточных организмов реальный предел мог бы иметь гораздо большее значение. Одноклеточные, живущие на больших глубинах, не погибают, когда их поднимают на поверхность, как и обитатели поверхности при их погружении вглубь. Многоклеточные же существа гибнут, особенно если переход происходит быстро. Предел солёности (табл. 2.3) следует оценивать по сравнению со средней солёностью морской воды, равной 2,95%. Интервал значений индекса рН простирается от сильно кислых до сильно щелочных показателей. В табл. 2.3 не вошло такое экстремальное воздействие, как радиация, в частности УФ излучение, заряженные частицы высоких энергий и гамма-излучение. Все они вызывают повреждения ДНК (разд. 2.4.4). Некоторые прокариоты могут устоять при таких уровнях облучения, которые нас с вами быстро отправили бы на тот свет; они даже способны существовать внутри ядерных реакторов! Экстремальные условия существуют в экстремальных местах обитания. Рассмотрим две такие среды, представляющие особый интерес с точки зрения поисков внеземной жизни.
Гидротермальные источники Гидротермальные источники, иногда называемые также чёрными курильщиками, обнаружены в зонах, где на границах литосферных плит происходит формирование новой океанической коры (см. разд. 2.1.2). Это своего рода башни, обычно имеющие метр в поперечнике и несколько метров в высоту, из которых изливается вода с температурой до 400 °С. В основном это та же океанская вода, просочившаяся в породы земной коры и вернувшаяся в океанический бассейн. В ней растворены такие газы, как СО2, Н2, СН4 и Н2S. Там, где эта горячая вода встречается с холодной и насыщенной кислородом океанской водой, происходят химические реакции с образованием твёрдых сульфидов, которые окрашивают воду в чёрный цвет и создают постройки типа упомянутых выше башен.
Имс 2.11 На рис. 2.11 приведена фотография такого источника, а на рис. 2.12 – некоторые из живых организмов, обнаруженных в его окрестностях.
Рис 2 12 Живые существа, заметные на рис. 2.12, - это трубчатые черви и паукообразные крабы. Они, а также двустворчатые моллюски встречаются здесь вместе с одноклеточными организмами. На подобных глубинах все эти существа являются пьезофилами. Вблизи жерла источника обитают гипертермофилы; к ним относятся прокариоты, многие из которых являются также анаэробами, поскольку вода гидротермальных источников почти лишена кислорода. Дальше от жерла температура падает примерно до 3 °С, и там обитают психрофилы. Таким образом, на расстояниях в несколько десятков метров мы переходим от организмов, приспособленных к жизни в горячей воде, к организмам, адаптированным к холодной среде обитания. Здесь и повсюду на больших океанских глубинах запасы пищи для гетеротрофных организмов обеспечивают минеральные и органические частицы, оседающие из приповерхностных вод, а дополнительным источником служат автотрофные прокариоты, которые, отмирая, поступают в пищевую цепь. Солнечный свет совершенно не проникает на такие глубины, и поэтому там не может происходить фотосинтез, вместо которого они осуществляют хемосинтез. Однако не совсем ясно, протекает ли хемосинтез полностью независимо от фотосинтеза. Например, метаногены используют СО2 и Н2, растворённые в воде, которая изливается из жерл гидротермальных источников – реакция (2.9). СО2 частично образуется при разложении карбонатов, погрузившихся в глубины земных недр, а частично – при окислении углерода, входящего в состав пород земной коры и мантии. Источником карбонатов в большой степени являются и раковины морских животных, в пищевой рацион которых входят организмы, осуществляющие фотосинтез. Кроме того, окисление углерода в какой-то мере может вызываться кислородом, содержащимся в воде океанов, который, в свою очередь, возникает в результате кислородного фотосинтеза – реакция (2.6). Биологическая составляющая может присутствовать и в Н2, хотя в большинстве случаев он происходит из небиологических источников, например путём окисления водой железа, входящего в состав новообразованных пород земной коры. Если хемосинтез в гидротермальных источниках не является независимым от остальной биосферы, где ещё мы могли бы найти организмы, осуществляющие его независимо? Один из возможных вариантов – породы земной коры.
Породы земной коры
Наверное, самым удивительным местом обитания живых организмов на Земле являются твёрдые среды – лёд и горные породы. В холодных пустынях Антарктиды, скажем в Сухих долинах Земли Королевы Виктории, встречаются обнажения выходящих на поверхность песчаников и кварцитов, в которых обитают колонии разнообразных одноклеточных организмов. Все они существуют в пределах сантиметра от поверхности – на такую глубину может проникать солнечный свет, а следовательно, здесь возможен фотосинтез. Несмотря на засушливый климат, в порах песчаника, как в ловушках, присутствует вода. Кроме того, хотя температура окружающего воздуха, как правило, ниже 0 °С, на глубине всего нескольких миллиметров от поверхности она уже может доходить до 10 °С. Стало быть, мы здесь встречаемся с не такими уж необычными условиями обитания живых существ; удивительно то, что эти условия обнаружены в горных породах в окружении гораздо более экстремальных сред. Ещё поразительнее присутствие в толще антарктического льда на глубинах до 2,5 км одноклеточных организмов, находящихся в состоянии анабиоза. Больше всего изумляют одноклеточные в породах земной коры на глубинах до нескольких километров. В настоящее время основная доля биомассы приходится на одноклеточные организмы, главным образом прокариоты, встречающиеся на значительной глубине под землёй. Заполненные водой поровые пространства и трещины в горных породах имеют достаточные размеры, и в них могут существовать одиночные клетки. Поры не изолированы друг от друга, так что земная кора больше напоминает губку, пронизанную одноклеточными организмами и окружающей их водной средой. В воде содержатся химические реагенты и происходят осуществляемые автотрофами разнообразные процессы хемосинтеза, в том числе метаногенез. Ферменты активизируют окислительно-восстановительные реакции, служащие главным источником энергии для жизненных процессов. Присутствие ферментов действительно необходимо – в противном случае имеющаяся химическая энергия высвобождалась бы спонтанно и быстро в результате небиологических процессов, и для живых организмов её вскоре не осталось бы. Небиологические процессы преобладают только при таких высоких температурах, при которых жизнь невозможна. В некоторых местах глубоко в земной коре осуществляющие хемосинтез автотрофы могут быть единственной основой пищевой цепи, и используемые ими для накопления энергии реагенты не зависят от остальной части биосферы. Действительно, живые организмы на Земле присутствуют, по-видимому, везде, где только может существовать жизнь, основанная на углероде и жидкой воде, т.е. в диапазоне температур от –18 до 123 °С в тех местах, где имеется вода в жидкой фазе и есть подходящий источник энергии.
2.6. Древо жизни
Живущие на Земле разнообразные организмы уже давно систематизированы, т.е. распределены по иерархической схеме, содержащей множество уровней. В мире растений и животных нижний уровень, который нас будет интересовать, - это вид. Два растения или пара животных относятся к разным видам, если половозрелая мужская особь одного вида и половозрелая женская особь другого не могут в естественных условиях произвести на свет способное к размножению потомство или же порождают такой гибрид крайне редко. На практике часто это бывает трудно проверить и вообще невозможно в отношении окаменелостей. Поэтому большинство организмов классифицируют исходя из их внешнего вида и поведения. Виды объединяют в роды, которые объединяют в cемейства, а следующие уровни иерархии называются отряд (порядок у растений), класс, тип (отдел у растений) и царство. Например, домашняя кошка (все разновидности) классифицируется таким образом: вид Catus (домашняя кошка), род Felis (дикие и домашние кошки), семейство Felidae (все кошачьи), отряд Carnivora (плотоядные), класс Mammalia (млекопитающие), тип Chordata (хордовые), царство Animalia (животные). Тип определяет характерное строение тела. Хордовые обладают спинным хребтом, и к ним относятся рыбы, лягушки и люди, но не пауки. Царство животных включает 24 типа. Начиная с 1970-х годов появилась возможность измерять биологическое различие (расстояние) между видами путём сравнения их РНК и ДНК. В результате мы кое-что узнаём и об эволюции. Например, два вида А и В, лишь немного отличающиеся друг от друга по составу своих нуклеиновых кислот, отклонились от общего предка, вероятно, позднее, чем виды А и С, проявляющие большее различие. С появлением этого метода в классификацию живых организмов были внесены фундаментальные изменения.
Рис 2.13 Результат приведен на рис. 2.13 – обычно его называют древом жизни. Самый верхний уровень иерархии называют доменом. На рисунке показаны три домена: Bacteria (бактерии), Archaea (археи) и Eukarya (эукариоты), плюс некоторые подразделы, входящие в состав царств и типов. Конец ветви может соответствовать царству (или типу), существующему в наши дни либо вымершему, эволюция которого никуда не привела. Вдали от концов находятся ныне исчезнувшие предки тех форм жизни, которые располагаются на концах. В точке ветвления лежит общий предок всех тех организмов, которые размещаются на ветвях, отходящих от этой точки. Дистанции вдоль ветвей характеризуют эволюционные расстояния, полученные измерениями сходств и различий нуклеиновых кислот у разных организмов. Таким образом, животные с растениями оказываются гораздо более близкими родственниками, чем те или другие - с метаногенами. Внутри каждого домена есть автотрофы и гетеротрофы. Eukarya включают в себя все организмы с эукариотическими клетками. На следующем уровне иерархии этот домен подразделяется на четыре царства, и вы наверняка знакомы с представителями царств Animalia (животные), Plantae (растения) и Fungi (например, дрожжи и грибы). Почти все члены этих трёх царств являются многоклеточными организмами, хотя некоторые низшие грибы – одноклеточные. Четвёртое царство Protista (простейшие) состоит в основном из
|
||||||||
|
|
Последнее изменение этой страницы: 2016-04-23; просмотров: 1179; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.108 (0.012 с.) |