Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Ранние свидетельства о живых организмахСодержание книги
Поиск на нашем сайте
Возраст самых древних горных пород, сохранившихся на Земле до наших дней, составляет примерно 3,85 млрд. лет, и этим же временем датируются первые свидетельства, имеющие отношение к жизни. Хотя в древнейших породах не обнаружено ископаемых остатков, в них имеются химические «следы» в форме отношений изотопов углерода. В ядре атома обычного изотопа углерода 12С содержится 6 протонов и 6 нейтронов, т.е. всего 12 нуклонов (протонов плюс нейтронов). В ядре менее широко распространённого изотопа 13С имеется 7 нейтронов. Обычная среда будет характеризоваться определённой величиной отношения n (12C)/ n (13C) числа ядер 12С и 13С, но если углерод участвовал в процессе фотосинтеза, то это отношение будет у него несколько более высоким. В небольших углеродных залежах, обнаруженных в древних породах, это отношение имеет повышенные значения по сравнению с его величиной в карбонатных породах, и лучше всего это можно объяснить тем, что тогда уже существовали живые организмы, способные к фотосинтезу. Древнейшие породы с такими «отметинами» обнаружены в комплексе Исуа на западе Гренландии. Однако есть и небиологические пути повышения изотопного отношения, и поэтому биологическая интерпретация представляется не бесспорной.
Рис 3.4(а)
Рис 3.4(б) Некоторые из древнейших ископаемых наблюдаются в форме строматолитов. На рис. 3.4(а) приведён пример современного строматолита. Эта структура состоит из слоёв минерального состава, отложенных колониями определённых бактерий, способных к кислородному фотосинтезу и называемых цианобактериями; эти организмы с успехом обитали в мелководной среде в довольно ограниченном диапазоне условий. На рис. 3.4(б) представлен ископаемый строматолит, образовавшийся, по-видимому, аналогичным путём. Древнейшие ископаемые остатки строматолитов имеют возраст 3,46 млрд. лет и обнаружены в Варравуна в Западной Австралии. Помимо поверхностного сходства с современными строматолитами они также содержат структуры, напоминающие окаменевшие бактерии. Последние вполне могли осуществлять фотосинтез, хотя и могли не выделять кислород. Тем не менее, интерпретация этих древних строматолитов как окаменелостей не является абсолютно надёжной, так как возможно их формирование и небиологическим путём. Признаки существования жизни в период 3,3 – 3,5 млрд. лет назад найдены также в зеленокаменном поясе Бабертон в Южной Африке, и среди них есть предполагаемые остатки термофильных прокариотов. Однако самые ранние свидетельства о наличии живых организмов, по которым практически не ведётся споров, – это ископаемые цианобактерии из Пилбары (Австралия) возрастом 2,7 млрд. лет и строматолиты, датируемые тем же временем, из Фортескью (тоже в Австралии).
Ранний фотосинтез
Первоначальные формы жизни существовали на Земле, которая сильно отличалась от нашей сегодняшней планеты. Одним из важнейших отличий был состав атмосферы. Атмосфера (и океаны) Земли сформировалась из газов, выделявшихся из горных пород и определивших её основной состав, а также летучих веществ, которыми были богаты планетезимали, бомбардировавшие молодую Землю (разд. 1.2.3). Среди летучих, достигших земной поверхности, должны были преобладать вода, СО2 и N2, а кроме того присутствовали в заметной концентрации другие вещества – СО и, возможно, СН4. Взаимодействуя с породами поверхности, они подвергались некоторой переработке, а фотодиссоциация воды под действием ультрафиолетового излучения Солнца привела к образованию кислорода (разд. 3.2.3), но в целом состав атмосферы мало изменился. Первичная атмосфера Земли имела слабо восстановительный характер, т.е. содержала молекулы, способные не приобретать электроны (как О2), а отдавать их (как СО и СН4). Если данные по изотопам углерода правильно интерпретированы, то это значит, что фотосинтез начался 3,85 млрд. лет назад. Но был ли он кислородным? Это маловероятно, хотя если основываться на полосчатых железистых кварцитах, то можно предположить и обратное. Эти породы осадочного происхождения состоят из слоёв, обогащённых оксидами железа и перемежающихся с обеднёнными железом породами при толщине каждого из слоёв порядка нескольких миллиметров. Они формировались в эти давние времена вплоть до примерно 2,0 млрд. лет назад. Как именно они возникали, в точности не известно, но не исключено, к примеру, что выделявшийся при фотосинтезе атмосферный кислород, растворённый в поверхностных водах, окислял соединения железа, поступавшие из жерл гидротермальных источников (разд. 2.5.2). Неясно, должен ли был для появления О2 происходить фотосинтез, по крайней мере, в самом начале, или хватило бы других источников, таких как фотодиссоциация воды. Кроме того, как следует из биологических данных, кислородный фотосинтез развился из комбинации двух некислородных форм, и считается маловероятным, что он мог осуществляться на столь ранней стадии. Лучшие свидетельства в пользу кислородного фотосинтеза в эпоху около 3,5 млрд. лет назад основаны на датированных этим временем ископаемых остатках строматолитов, если они являются продуктами жизнедеятельности цианобактерий.
Последующие события
Только примерно к 2,3 млрд. лет назад содержание О2 в атмосфере превысило следовое количество. О возможных причинах будет сказано в разд. 3.2.3. К этому времени уже вполне утвердились эукариоты, а первые эукариотические клетки или их ныне вымершие предки появились около 2,7 млрд. лет назад. Все прежде существовавшие организмы были прокариотами. Появление эукариотов стало одним из крупнейших достижений в развитии жизни на Земле, и с того времени они начали быстро эволюционировать. Сложная структура эукариотической клетки, по-видимому, возникла благодаря симбиозу, в котором объединились две или более прокариотические клетки. Об этом свидетельствуют некоторые органеллы в эукариотических клетках (разд. 2.3). Например, хлоропласты растительных клеток, в которых осуществляется фотосинтез, и митохондрии, обеспечивающие дыхание у растительных и животных клеток, обладают ДНК. Это свидетельствует о том, что их ближайшими родственниками являются бактерии. Среди объединившихся прокариотов есть как анаэробные, так и аэробные формы; следовательно, многие эукариотические клетки были способны использовать кислород для аэробного дыхания. Сегодня, почти без исключений, эукариотическим клеткам требуется кислород, который либо присутствует в воздухе, либо растворён в воде, тогда как одних прокариотов кислород убивает, другие могут его выносить, и лишь небольшой их доле он действительно нужен. Почти все многоклеточные организмы состоят из эукариотических клеток, и их широкое распространение явилось следующим крупным событием, которое можно отметить на временной оси развития жизни на Земле. Оно произошло довольно поздно – примерно 610 млн. лет назад, когда прежде весьма редко встречавшиеся многоклеточные организмы стали встречаться гораздо чаще, приобрели более крупные размеры и более сложное строение. Появились кораллы и существа, похожие на медуз, а также какие-то «прошитые» создания, не имеющие современных аналогов. «Вскоре» после этого, 545 млн. лет назад, в огромной степени возросло количество таких организмов, возникло множество новых видов и, кроме того, расширился диапазон их сред обитания. Этот довольно точный момент времени фиксирует начало кембрийского периода (545 – 495 млн. лет назад) в геологической истории. Многие из новых организмов содержали такие твёрдые части, как раковины и наружные скелеты, и поэтому они смогли сохраниться в виде окаменелостей.
Рис 3.5 На рис. 3.5 показан ископаемый остаток одного из многочисленных видов трилобитов, которые в изобилии обитали в океанах начиная с раннекембрийского времени. Хотя в ту эпоху происходили изменения в окружающей среде, не ясно, они ли были причиной кембрийской «вспышки» этих животных. Дополнительную роль могли сыграть рост содержания О2 в атмосфере (разд. 3.2.3) и возникновение новых разнообразных сред обитания в результате континентального дрейфа, вызванного тектоникой плит. До наступления кембрийского периода почти вся жизнь была сосредоточена в океанах. На более ранних этапах, даже 2,7 млрд. лет назад, на суше встречались отдельные колонии бактерий, но очень редко. И только в кембрии более разнообразные формы жизни сумели гораздо шире распространиться на суше. Это началось примерно 490 млн. лет назад, т.е. в конце кембрийского периода, а тотальное освоение суши происходило между 440 и 420 млн. лет назад. К моменту 420 млн. лет от настоящего времени все основные группы многоклеточных организмов – беспозвоночные, позвоночные и растения - обитали на суше и в море. Род человеческий (группа родственных видов) Homo появился всего 3 млн. лет назад, а вид Homo sapiens, к которому мы с вами принадлежим, – вообще лишь 0,1 млн. лет назад!
Массовые вымирания
Два самых драматичных массовых исчезновения живых организмов произошли 248 млн. лет назад, в конце пермского периода, и 65 млн. лет назад, в конце мелового периода (рис. 3.2). Более позднее из них определяет конец мезозойской эры («эры средней жизни»), когда вымерло 70% морских видов плюс большая доля наземных организмов, в том числе, как известно, динозавры. Более раннее массовое вымирание соответствует концу палеозойской эры («эры древней жизни»); тогда оказалось потеряно до 95% морских видов плюс высокий процент видов, обитавших на суше. В каждом случае для исчезновения всех видов потребовалось 0,1–1 млн. лет, т.е. массовое вымирание – вовсе не мгновенное явление. Восстановление биосферы заняло в десять раз больше времени. Произошло ещё три менее массовых вымирания: около 450, 360 и 210 млн. лет назад и, по всей вероятности, ещё несколько в более древние времена. Свидетельства более ранних событий менее надёжны, в какой-то степени из-за менее полной палеонтологической летописи, дошедшей до нашего времени.
3.2.2. Причины массовых вымираний
Чтобы найти причину массового исчезновения живых организмов, нужно поискать какое-то глобальное изменение в окружающей среде, иначе многие виды могли бы мигрировать туда, где среда обитания была для них подходящей. К тому же это изменение должно быть внезапным – в противном случае многие виды приспособились бы к новым условиям в ходе нормального эволюционного развития. Сейчас получены важные свидетельства того, что массовое вымирание 65 млн. лет назад было вызвано столкновением Земли с кометой или астероидом. В море вблизи северного побережья мексиканского полуострова Юкатан были обнаружены остатки крупного ударного кратера Чиксулуб, возраст которого определён в 65,5 млн. лет. Его могло образовать небесное тело диаметром около 10 км, двигавшееся со скоростью 30 км/с. В результате такого столкновения должно было произойти изменение климата, в частности, из-за появления большого количества пыли в верхней атмосфере, вызвавшей глобальное похолодание. Пыль непосредственно возникла при ударе тела о Землю, а также от частиц дыма, сопровождавшего лесные пожары, спровоцированные тепловым ударом. Выделенной при столкновении энергии было вполне достаточно, чтобы привести к массовому вымиранию. Образовавшаяся пыль распределилась по всему земному шару, о чём свидетельствует аномально высокая концентрация иридия в осадочных породах возрастом 65 млн. лет, обнаруженная во многих странах. Иридий в гораздо большем количестве присутствует в метеоритах, чем в земной коре. В сущности, именно обнаружение этого обогащённого иридием слоя и привело к открытию кратера Чиксулуб. Хотя столкновение с космическим телом и сыграло роль в массовом вымирании 65 млн. лет назад, оно, конечно, было не единственным фактором. Некоторые виды уже находились в состоянии упадка, и скорость их исчезновения возрастала ещё до удара. Это могло быть следствием уже происходившего изменения климата, в частности, глобального похолодания и связанного с ним понижения уровня моря. К изменению климата могли привести многие причины; так, для данного конкретного похолодания мог иметь значение выброс пыли в атмосферу в результате крупных вулканических извержений в Индии около 65 млн. лет назад. Согласно другой гипотезе, похолоданию могло бы способствовать уменьшение вулканической активности на планете, поскольку из-за этого выделялось бы меньше парникового газа СО2. Таким образом, хотя уменьшение численности и исчезновение одних биологических видов было спровоцировано столкновением с космическим телом, для других это просто стало последней каплей. Третьи же сохранились в целости и невредимости; среди них – акулы, многие пресмыкающиеся и предки современных млекопитающих. Массовое вымирание, произошедшее 248 млн. лет назад, понять труднее. Как известно, в то время на обширных пространствах Сибири происходили сильные извержения вулканов. Кроме того, процессы тектоники плит вели к изменениям очертаний континентов и океанических бассейнов, переменам в системах океанических течений, понижению уровня моря и частичному удалению мелководного шельфа. Есть также признаки некоторого уменьшения содержания О2 в атмосфере. Согласно одному из возможных сценариев, вначале сибирские вулканические извержения привели, вследствие большого количества попавшей в атмосферу пыли, к продолжавшемуся несколько лет глобальному похолоданию, а после того как пыль осела, из-за роста СО2 началось глобальное потепление. Средняя глобальная температура поверхности могла подняться на 5 °С. Погибли многие виды наземных организмов. Постепенно разогрелись океаны, что вызвало вымирание многих биологических видов и в этой среде обитания. Затем рост температуры океана привёл к выделению из донных осадков СН4 – сильного парникового газа, который мог стать причиной дальнейшего повышения средней температуры ещё на 5 °С, что привело к ещё более крупным вымираниям. Три других массовых вымирания, начиная с кембрийского периода, ассоциируются с климатическими изменениями, в частности – с наступлением ледниковых эпох, хотя причины этих перемен не вполне ясны. Если массовые вымирания происходили также и задолго до 248 млн. лет назад, – а это вполне вероятно, – то об их причинах мы можем только гадать. Могли сыграть роль как те факторы, о которых говорилось выше, так и некоторые другие. Например, массовое исчезновение живых организмов могло произойти, когда после момента времени 2,5 млрд. лет назад начало возрастать содержание кислорода в атмосфере: в то время для большинства видов он должен был быть губителен. Даже в наши дни кислород ядовит для многих анаэробных прокариотов. Ещё одна причина – взрыв близкой сверхновой. Этот вопрос рассматривается в гл. 7: сверхновые могли играть роль в среднем каждые несколько сотен миллионов лет. В целом массовые вымирания, вероятно, способствовали увеличению разнообразия видов и усложнению структуры организмов. Возможно, это было не так уж плохо. А если бы не было планеты-гиганта Юпитера, то столкновения комет с Землёй были бы более частыми, ведь Юпитер играет роль своеобразного гравитационного экрана. Это могло бы привести к стерилизации нашей планеты, погасило бы процесс зарождения жизни или задержало появление многоклеточных организмов.
3.2.3. Воздействие биосферы на атмосферу Земли
Современный состав атмосферы указан в табл. 2.1. Основные её компоненты – это N2 и О2. А в то время, когда на Земле обитал последний общий предок, в воздухе преобладали СО2 и N2, в значительных количествах присутствовало также несколько других газов, например СО, и почти не было О2, хотя вода (в виде пара) в атмосфере имелась всегда.
Кислород
Один из путей получения О2 – фотодиссоциация (фотолиз) водяного пара под действием солнечного ультрафиолетового излучения. Процесс начинается с разложения молекулы Н2О под действием кванта УФ излучения:
Н2О ® Н + ОН. (3.1)
Затем проходит ещё несколько стадий, так что весь процесс можно описать в виде следующей химической реакции:
2Н2О ® 2Н2 + О2. (3.2)
В результате О2 будет накапливаться, поскольку Н2, с которым кислород мог бы вступить в реакцию, улетучивается в космическое пространство. Однако гораздо более обильным источником О2 служит кислородный фотосинтез, при котором молекулярный кислород выделяется как побочный продукт (разд. 2.4.2); именно благодаря этому процессу О2 стал одной из важнейших составляющих атмосферы. Заметим, что кислородный фотосинтез мог бы возникнуть примерно 3,5 млрд. лет назад, но уже наверняка он происходил благодаря цианобактериям 2,7 млрд. лет назад. Однако из геологических данных, касающихся горных пород, возраст которых установлен, и изотопных отношений следует, что приблизительно 2,3 млрд. лет назад атмосфера не носила окислительного характера, а после этого времени уже была окислительной. Эта перемена объяснялась ростом содержания кислорода, очевидно, из-за изменения в характере квазиравновесия между скоростью образования О2 и скоростью его исчезновения. В течение всех последних 2,3 млрд. лет скорость генерации кислорода в процессе фотосинтеза, по-видимому, была практически постоянной, и в таком случае скорость его удаления должна была идти на убыль. К потере О2 ведёт окисление органических соединений в процессе дыхания (разд. 2.4.2) и в ходе гниения отмерших организмов. Его количество уменьшается и при окислении слабоокисленных пород и газов. Согласно одной из гипотез, в результате разложения анаэробами органического вещества, образовавшегося в ходе фотосинтеза, степень окисленности пород земной коры возрастает, а следовательно, уменьшается интенсивность поглощения ими атмосферного О2. При анаэробном разложении образуются О2 и СН4, и если метан уходит в космическое пространство (или, по крайней мере, уходит входящий в него Н2), то в результате степень окисленности пород увеличится. Тем не менее причина возрастания О2 в атмосфере нам ещё далеко не ясна.
Рис 3.6 Процессы установления квазиравновесия на протяжении истории Земли привели к тому, что в течение большей части времени содержание О2 в атмосфере менялось – одна из возможных интерпретаций довольно ограниченного количества данных представлена на рис. 3.6. Если бы не происходил кислородный фотосинтез, содержание молекулярного кислорода в атмосфере всегда было бы значительно ниже чем 1%. Большая распространённость О2 приводит к очень низкому содержанию веществ, обладающих восстановительными свойствами, например СО и СН4. Заметим, что СН4 очень легко окисляется, и поэтому если бы метан не возникал в изобилии в результате биосферных процессов под действием метаногенов (разд. 2.4.2), то его содержалось бы в атмосфере ещё меньше, чем сейчас, когда его относительная концентрация равна 1,6 ´ 10–6. В океанах содержание О2 тоже изменчиво. Верхние несколько сотен метров тесно контактируют с атмосферой, и в них содержание кислорода близко к атмосферному. Более глубокие слои всегда были обеднены О2, и аэробное дыхание должно было стать возможным только после заметного увеличения его содержания в атмосфере. Оценки содержания кислорода в глубоких слоях океана имеются только вплоть до 1 млрд. лет назад.
Озон (О3)
В разд. 2.1.3 отмечалось, что О3 защищает всё живое от разрушающего воздействия УФ излучения Солнца. При этом поглощается большая доля этого излучения. Озон образуется из О2 под действием солнечных УФ фотонов, и поэтому изменение его содержания в функции времени соответствует характеру изменения О2. Однако достаточное для создания защитного щита количество О3 обеспечивается довольно небольшим количеством О2. Следовательно, начиная с самых первых дней существования на Земле биосферы, в атмосфере содержалось достаточно О2, чтобы защитить живые организмы от солнечного ультрафиолета.
Диоксид углерода
Потеря большого количества СО2 обусловлена процессами в биосфере. Наибольшая его доля связана в горных породах, особенно в карбонатных минералах (например, кальците СaCO3). Именно из этих минералов в основном состоят различные типы известняка, в том числе и знакомый всем писчий мел. Известняк – это осадочная порода, которая при длительном воздействии высокой температуры и давления превращается в метаморфическую породу типа мрамора. Большинство карбонатных пород образуются на океаническом дне, связывая СО2, растворённый в морской воде. На место израсходованного диоксида углерода поступает новый из атмосферы, где его количество при этом должно было бы уменьшаться. Но в отсутствие живых организмов этот процесс происходил очень медленно, поэтому, скорее всего, концентрация СО2 в атмосфере поддерживалась гораздо большей, чем сейчас, за счёт его поступления от вулканических извержений и эрозии горных пород. Биосфера способствует исчезновению углекислого газа, который идёт на формирование скелетов морских животных и образование органических соединений. Он лишь частично возвращается в атмосферу благодаря дыханию и процессам распада при захоронении части органогенного углерода, когда образуются разнообразные осадочные отложения, в том числе горючие полезные ископаемые. Хотя сейчас в атмосфере присутствуют лишь следы СО2, этот газ играет немалую роль, так как является главной причиной парникового эффекта, другим важным фактором в создании которого служит водяной пар (разд. 2.1.3). Не будь всеобъемлющего парникового эффекта, средняя глобальная температура земной поверхности была бы примерно на 33 °С ниже, чем теперь. Благодаря присутствию в атмосфере следов СО2 происходит фотосинтез, и продолжительное время существуют зелёные растения, водоросли, некоторые прокариоты и все зависящие от них формы жизни. Кроме того, СО2 помогает решить ещё одну проблему. Дело в следующем. Согласно оценкам, в самое раннее время, когда мог существовать последний общий предок, т.е. чуть раньше чем 3,85 млрд. лет назад, наше Солнце грело на 25% слабее, чем сейчас. Это приводит нас к проблеме тусклого Солнца. Ведь если бы у Земли тогда была такая атмосфера, как теперь, земная поверхность, возможно, находилась бы в замёрзшем состоянии, а восстановиться из этого состояния было бы трудно. Однако, исходя из геологических данных, вода в жидкой фазе в то время и позднее была широко распространена на планете. Одно из решений этой проблемы даёт динамика карбонатно-силикатного цикла. Допустим, что поверхность Земли охлаждается. Вследствие этого уменьшается интенсивность эрозии силикатов под действием СО2 и Н2О и, следовательно, скорость формирования карбонатов. Стало быть, снижается скорость удаления СО2 из атмосферы. Диоксид углерода выделяется в атмосферу при извержениях вулканов, и если он расходуется медленнее, содержание его в воздухе повышается. Это приводит к усилению парникового эффекта, и земная поверхность разогревается. В результате в период тусклого Солнца Земля могла бы не замёрзнуть. В более длительной перспективе, когда светимость Солнца возрастала, скорость эрозии увеличивалась и, соответственно, содержание СО2 в атмосфере падало, препятствуя таким образом сильному увеличению температуры. Это добавлялось к потерям СО2 вследствие биосферных процессов. Альтернативное решение проблемы тусклого Солнца дает СН4. Этот газ создаёт столь сильный парниковый эффект, что даже мизерное его количество в атмосфере могло бы помешать замерзанию поверхности. Метана требуется больше, чем присутствует в атмосфере в настоящее время; но поскольку кислорода тогда было крайне мало, достаточное количество СН4 могло сохраняться.
Азот Под действием биосферы содержание СО2 в атмосфере уменьшается, а на самую обильную составляющую воздуха – N2 – живые организмы оказывают противоположное влияние. Азот выводится из атмосферы при электрических разрядах, в частности молниях, когда образуются оксиды азота, вымываемые дождём. В отсутствие биосферы N2 может возвращаться в атмосферу только в результате медленно протекающих процессов эрозии пород земной коры. Что касается живых организмов, то у одних прокариотов из органического вещества азот выделяется и, хотя другие поглощают N2, суммарным эффектом является рост интенсивности его выделения. В результате, поддерживается более высокое содержание азота, чем было бы на стерильной планете. Следовательно, современная атмосфера Земли в очень большой степени является продуктом её биосферы.
3.2.4. Определение абсолютного возраста
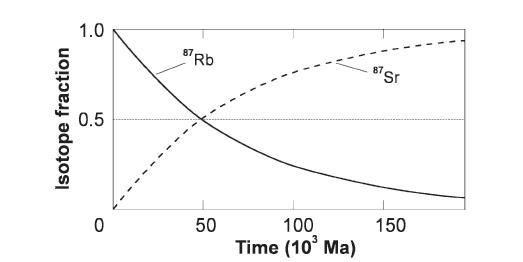
Рис 3.7 Время, когда в истории Земли произошли те или иные события, определяется методами радиоизотопного датирования. Рассмотрим, например, рубидиево-стронциевый метод. Предположим, что происходит затвердевание горной породы и что она содержит минерал, включающий в себя такой элемент, как рубидий. Некоторая доля этого рубидия представлена нестабильным изотопом 87Rb, при радиоактивном распаде которого образуется стабильный изотоп стронция 87Sr (рис. 3.7). Допустим, что в затвердевавшей породе не было 87Sr и что она не теряла ни 87Rb, ни рождающийся из него 87Sr. Тогда отношение числа ядер этих изотопов n (87Sr)/ n (87Rb) будет постепенно возрастать со временем. Пусть мы знаем скорость, с которой 87Rb распадается и превращается в 87Sr. Тогда, если в некоторый момент времени t мы измерим величину указанного отношения ядер, мы сможем сказать, сколько времени прошло после затвердевания породы. Скорость превращения выражается через период полураспада – время, за которое распадается половина всех ядер. Для рассматриваемого нами процесса период полураспада равен 48,8 млрд. лет. Считая, что в момент затвердевания (t = 0) отношение n (87Sr)/ n (87Rb) = 0, получаем, что в момент t = 48,8 млрд. лет это отношение равно 1, в момент t = 2 ´ 48,8 млрд. лет – 3, и т.д. Точность этого метода ограничивается тем обстоятельством, что в момент затвердевания породы в ней всё же могли присутствовать оба изотопа. Правда, об отношении числа их атомов в момент t = 0, при некоторых допущениях, можно судить по относительному обилию других изотопов, содержащихся в породе. Возраст затвердевания важен потому, что он указывает нам время, когда порода сформировалась. Горные породы, образующиеся при затвердевании расплавленной магмы, называются изверженными, или магматическими. Однако в породах, испытавших плавление, не могут сохраняться ископаемые остатки живых организмов. Поэтому для того, чтобы определить возраст событий, рассказывающих нам о развитии биосферы, используются магматические породы, вмещающие в себя породы другого типа, в которых ископаемые остатки сохраняются, а именно осадочные и некоторые метаморфические породы. Значения возраста изверженных пород заключают в себе возраст находящихся между ними осадочных пород. Датируя таким способом осадочные породы, мы можем также определить время других событий, например, роста содержания кислорода в атмосфере, так как осадочные слои, сформировавшиеся в океанах, обогащённых кислородом, отличаются от осадков, образовавшихся в обеднённых кислородом условиях. Имея столь долгий период полураспада, система изотопов Rb – Sr позволяет определять возраст событий вплоть до возникновения Солнечной системы. Другие пары изотопов также имеют большие периоды полураспада и используются для датировки самых разнообразных промежуточных событий. Среди них - бомбардировка Земли космическими телами, происходившая ранее 3,8 млрд. лет назад: этим возрастом датируются древнейшие из сохранившихся земных пород (древнее них лишь несколько редких исключений). Историю этих событий удалось проследить благодаря изучению образцов лунных пород и определению их времени затвердевания после расплавления, вызванного ударами метеоритов. Оказалось, что почти вся поверхность нашего ближайшего космического соседа древнее 3,8 млрд. лет. Определив радиоизотопные возрасты веществ, входящих в состав большинства древнейших метеоритов, нашли, что возраст Земли, да и Солнечной системы в целом равен 4,6 млрд. лет. Для исследования событий недавнего времени, происходивших в последние, скажем, 100 тыс. лет, используются изотопы с более короткими периодами полураспада, для того чтобы успели возникнуть изменения, которые можно было бы измерить. Изотоп 14С имеет период полураспада всего 5730 лет и служит основой радиоуглеродного метода датирования. В состав атмосферного СО2 входит малая доля 14С, возникающего при взаимодействии космических лучей с 14N – основным изотопом азота. Живые организмы обмениваются углеродом с атмосферным воздухом в процессе фотосинтеза, во время еды и дыхания, поэтому отношение 14С к углероду в целом у них почти не отличается от того значения, которое существует в атмосфере. После смерти этот обмен сильно замедляется, потому отношение n (14C)/ n (C) начинает монотонно уменьшаться. Следовательно, сравнивая величины этого отношения в мёртвом веществе и в живой биосфере, можно определить время отмирания органического вещества. Точность получаемых результатов снижается из-за многих неопределённостей (например, значение этого отношения для атмосферы в прошлом). В результате для эпохи 20 000 лет назад погрешность составляет около 3000 лет. С продвижением в глубь веков точность ухудшается, так что предел для определения возраста радиоуглеродным методом составляет примерно 70 000 лет.
3.3. Происхождение жизни на Земле
Как говорилось в начале этой главы, последним общим предком была прокариотическая клетка какого-то примитивного вида. Носителем её генетической информации, конечно, являлась ДНК, а белки были основой для выполнения многочисленных функций, осуществляемых и сегодня белками. Но эта клетка не могла быть первой формой жизни на Земле, появившейся непосредственно из относительно малых органических молекул. Это был бы слишком уж большой скачок. Основная трудность состоит в проблеме «курицы и яйца». Для синтеза белков из малых молекул требуется генетическая информация, содержащаяся в ДНК, а чтобы этот синтез мог происходить, в качестве ферментов нужны белки (разд. 2.4.3). Далее, белки как ферменты требуются для воспроизведения ДНК в процессе размножения (разд. 2.4.4). Таким образом, мы не можем иметь ДНК без белков и белки – без ДНК.
3.3.1. Мир РНК
Одно из решений этой дилеммы предложили в конце 1960-х годов независимо друг от друга несколько учёных. Предполагалось, что у предшественника последнего общего предка живых организмов носителем генетической информации служила не ДНК, а РНК. Решающим моментом здесь является то, что РНК в этой теории самовоспроизводится без помощи белков и служит катализатором всех химических реакций, необходимых для выживания и репродуцирования этого предшественника, в том числе –для синтеза белков. Мир, в котором жизнь таким именно образом базируется на РНК, называют миром РНК. Основания для этой гипотезы были следующие: · нуклеотиды в составе РНК легче синтезируются, чем в ДНК; · легко видеть, как ДНК может развиться из РНК и затем, будучи более устойчивым образованием, принять на себя роль вместилища генетической информации (в последующие годы было обнаружено, что, когда ретровирусы поражают эукариотические клетки, они могут создавать ДНК на основе собственной РНК); · было трудно понять, как белки могли бы самовоспроизводиться в отсутствие нуклеиновых кислот (хотя недавно обнаружено, что некоторые белки способны к саморепликации). Однако в конце 1960-х годов концепция мира РНК действительно встретилась с большой трудностью, а именно: совсем не было известно, чтобы РНК действовала как катализатор для каких-либо биохимических реакций в современной биосфере. Затем в 1983 г. американский биохимик Томас Сеч и канадский биолог Сидни Альтман независимо друг от друга открыли ферменты (энзимы), состоящие из РНК, – рибозимы. Хотя эти рибозимы были способны делать немногим больше, чем разрывать и соединять существовавшие до этого РНК, учёные акцентировали внимание на том, что некоторые древние формы РНК могли делать гораздо больше. Кроме того, хотя некоторые стадии процесса репродукции РНК, в котором сама РНК служит катализатором, остаются весьма туманными, другие стадии были продемонстрированы лабораторными экспериментами. После 1983 г. также было установлено, что у рибосом (разд. 2.4.3), вероятно, именно РНК, а не белок, служит катализатором синтеза белка. И всё же, остаётся гигантская брешь между тем, что, как мы знаем, способна делать РНК, и тем, что ей необходимо делать в мире РНК. Если допустить, что мир РНК существовал, получается колоссальный скачок назад от последнего общего предка в направлении возникновения жизни. Но это ни в коей мере не определяет для нас весь путь – ведь для того, чтобы получить РНК, нам нужно иметь запас её составляющих – основания, сахар рибозу и фосфаты. Синтезировать их трудно, особенно некоторые из оснований и рибозу. Было поставлено множество экспериментов, в которых смеси газов, моделирующие земную атмосферу того времени со слабо восстановительными свойствами (а возможно, и нейтральными), подвергались воздействию имитаций солнечного излучения, молний и других источников энергии. Было получено много разных органических соединений, в том числе некоторое количество аминокислот и прочих малых органических молекул, имеющих отношение к жизни. К сожалению, другие важные молекулы образовались лишь в пренебрежимо малых долях, и среди них злополучные основания РНК и рибоза. Источниками некоторых недостающих компонентов (но не в достаточно больших количествах и, к тому же, вероятно, не всех необходимых) могли бы послужить гидротермальные системы с температурами немного выше 100 °С. Одним из возможных вариантов является поступление требуемых биомолекул из космоса. Падение обогащённых летучими веществами метеоритов и кометных осколков должно было обеспечить Землю значительными количествами того, что называют добиологическими молекулами. Предполагается, что они способствовали появлению на нашей планете жизни на основе углерода и жидкой воды. Метеориты, представляющие собой фрагменты астероидов, бывают нескольких типов, и в одном из них – так называемых углистых хондритах – обнаруживается множество различных органических соединений, включая аминокислоты. Изучение комет показывает, что и в них содержатся органические соединения. Однако основания и рибоза не наблюдаются, откуда следует, что в прошлом они могли отсутствовать. Но даже если бы у нас были все компоненты, остаётся ещё большая проблема, как собрать их, чтобы образовались нуклеотиды, и объединить нуклеотиды в нить РНК, содержащую порядка 105 оснований, необходимых в качестве фундамента жизни. Дополнительная трудность связана с вероятным обилием молекул, родственных нуклеотидам РНК. Включение их в РНК привело бы к нарушению её функций. Возможный способ решения этой проблемы рассмотрен в разд. 3.3.3.
3.3.2. Происхождение клетки
Раздумывая над происхождением биохимических компонентов жизни, нам нужно предусмотреть и то, чтобы они находились в защищённой среде – клетке. Каким образом появилась первая настоящая клетка, нам не известно, как и многое другое. Однако некоторой путеводной нитью могли бы послужить мелкие капли, называемые коацервата |
||||
|
|
Последнее изменение этой страницы: 2016-04-23; просмотров: 434; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.117.8.76 (0.021 с.) |