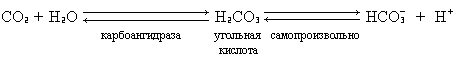Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Фракционное осаждение белков методом высаливанияСодержание книги
Поиск на нашем сайте
* Разделение альбуминов и глобулинов яичного белка с использованием сульфата аммония 1. К 1 мл неразбавленного яичного белка приливают 6 мл воды. 2. Для растворения образовавшегося небольшого белого хлопьевидного осадка глобулинов в эту пробирку приливают несколько капель насыщенного раствора сульфата аммония. 3. К 7 мл данного раствора яичного белка, приливают равный объем насыщенного раствора сульфата аммония. 4. Наблюдают выпадение осадка глобулинов, который удаляют фильтрованием. 5. К фильтрату добавляют кристаллический сульфат аммония до полного насыщения. 6. Выпавший при этом осадок альбуминов также отфильтровывают. 7. С фильтратом проделывают пробу на полноту осаждения белка с помощью 5% ТХУ.
* Обратимость высаливания 1. К 2 мл 1% раствора белка приливают 2 мл насыщенного раствора сульфата аммония. 2. Далее в пробирку наливают 4 мл воды и встряхивают. 3. Наблюдают, записывают и объясняют результаты.
* Разделение альбуминов и глобулинов яичного белка с использованием хлористого натрия и сернокислого магния 1. В две пробирки наливают по 3 мл белка. 2. Прибавляют до полного насыщения в одну пробирку тонко измельченный NaCl, в другую – MgSO4. 3. Отфильтровывают появившиеся через несколько минут осадки глобулинов. 4. К фильтратам прибавляют по 5 капель 1% раствора уксусной кислоты. 5. Наблюдают выпадение альбуминов, отфильтровывают осадок. 6. Фильтрат проверяют на отсутствие белка при помощи биуретовой реакции.
Оформление работы
К занятию: 1. Кратко законспектировать теоретические материалы к лабораторной работе. Во время занятия: 2. Описать этапы работы. 3. Описать результаты. 4. Сделать выводы.
Методические указания к лабораторному практикуму по курсу "Общая и экологическая биохимия" Раздел "Белки" Лабораторная работа № 3
Оборудование и материалы: · Спектрофотометр SOLAR · Центрифуга К-24 · Центрифуга настольная · Кюветы стекляные · Термостат · Баня песочная · Пипетки стекляные на 1 мл и 5 мл · Микропипетки автоматические · Цилиндры мерные на 250 мл и 100 мл · Колбы емкостью 250 мл и 100 мл · Пробирки · Штативы для пробирок · Бумага фильтровальная · Индикаторная бумага универсальная · Бумага миллиметровая
Реактивы: · Молоко · Кровь цельная · Биуретовый реактив · реактив Брэдфорд · раствор А (2%-ный Na2CO3 в 0,1М NaOH) · раствор Б (0,5%-ный CuSO4· 5Н2О в 1%-ном тартрате натрия (или калия) · коммерческий реактив Фолина · Бензидин, раствор · Перекись водорода (Н2О2), 3% раствор · Молибденовый реактив: молибдат аммония ([NH4]2MoO4), 15% раствор / HNO3(конц.) в отнош. 110/ 90 · Гидроксид натрия (NaOH), 5 М раствор · Карбонат натрия (Na2CO3), 0,1% раствор · Сульфат аммония ([NH4]2SO4), насыщ. раствор · Эфир диэтиловый · Спирт этиловый, 96% · Спирт метиловый · Хлороформ · Уксусная кислота, ледяная, 0,1% раствор · Азотная кислота (HNO3), конц. · Серная кислота (H2SO4), конц., 10 М раствор · стандартные растворы белка (БСА) №1, 2, 3 · Растворы белка разных концентраций (исследуемые образцы) · вода дистиллированная Теоретическая часть Сложные белки
Сложные белки состоят из полипептидных цепей, построенных из аминокислот, а также включают неаминокислотные компоненты, представленные кофакторами и простетическими группами. В их число входят порфирины, производные витаминов, липидные, углеводные, нуклеиновые и другие компоненты. Одной из наиболее изученных и интересных групп сложных белков являются гемопротеиды.
Гемопротеиды
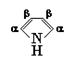
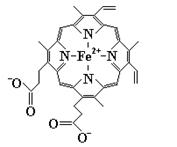
Гемсодержащие белки участвуют в процессах транспорта электронов, транспорта кислорода, в процессе фотосинтеза. Классическими представителями сложных белков этой группы являются миоглобин и гемоглобин. На их примере можно четко проследить связь между структурой и функцией глобулярных белков. Эти гемопротеиды содержат в качестве простетической группы гем – циклический тетрапиррол, включающий атом железа, присутствие которого объясняет красный цвет данных белков и их способность запасать кислород (миоглобин) или обеспечивать его транспорт (гемоглобин). Тетрапирролы состоят из четырех молекул пиррола (рис. 3.1), связанных между собой четырьмя a-метиновыми мостиками, что обеспечивает образование плоской кольцевой структуры, несущей в определенных положениях соответствующие заместители.
Рис. 3.1 Структурная формула пиррола. В тетрапиррольных кольцах могут присутствовать разные b-заместители – метильные (М), винильные (V) и пропионатные (Pr) группы. Например, в составе гема указанные заместители расположены в следующем порядке: M, V, M, V, M, Pr, Pr, M (рис. 3.2).
Рис. 3.2 Структура гема.
В центре плоского тетрапиррольного кольца находится один атом железа в ферро(Fe2+)-состоянии. Похожие по строению простетические группы, содержащие ионы металлов, обнаруживают и в других белках: цитохромах, хлорофилл-содержащих белках (Mg2+) и некоторых ферментах, например, каталазе, триптофанпирролазе и др. В цитохромах происходит попеременное окисление и восстановление атома железа (Fe2+ ↔ Fe3+), играющее определяющую роль в их функционировании. Напротив, окисление Fe2+ до Fe3+ в миоглобине или гемоглобине сопровождается потерей их биологической активности.
Гемоглобин
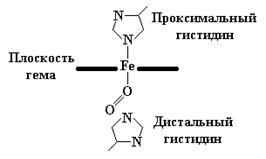
Гемоглобины – структурно-родственные белки эритроцитов позвоночных, выполняющие две важные биологические функции: 1) перенос О2 из легких к периферическим тканям; 2) перенос СО2 и протонов от периферических тканей к дыхательным органам для последующего выделения из организма. Гемоглобины представляют собой тетрамерные белки, молекулы которых образованы попарно ассоциированными разными типами полипептидных цепей, которые обозначаются буквами a, b, g, d, S и др. В состав тетрамерной молекулы входят по две цепи двух разных типов. Гемоглобин взрослого человека, обозначаемый А1, состоит из двух a- и двух b-цепей. Их длина примерно одинакова - a-цепь содержит 141 остаток, а b-цепь – 146. В молекуле гемоглобина гем расположен в щели между двумя a-спирализованными участками каждой цепи. Его пропионатные группы ориентированы к поверхности глобулы, а другие заместители обращены внутрь глобулы и окружены неполярными (гидрофобными) аминокислотными остатками. Исключение составляют два остатка гистидина – проксимальный и дистальный, которые расположены вблизи атома железа. Причем, атом азота гетероциклического кольца проксимального гистидина связан пятой координационной связью с атомом железа (четыре другие связи атом железа образует с азотами пиррольных колец гема). В норме дистальный гистидин не взаимодействует с атомом железа, который предназначен для связывания молекулы кислорода. При оксигенации гемоглобина образуется связь между атомом кислорода и атомом Fe2+, направленная перпендикулярно плоскости кольца гема. Второй атом кислорода удален от дистального гистидина, и связь между атомами кислорода образует относительно плоскости гема угол 121º (рис. 3.3).
Рис. 3.3 Ориентация молекулы кислорода, связанной с атомом железа гема в составе гемоглобина.
Цепи гемоглобина А1 – a и b кодируются разными генами и имеют разную первичную структуру. В тоже время первичная структура гомологичных b-, g- и d-цепей гемоглобина человека в значительной степени сходна. Свойства индивидуальных гемоглобинов определяются их олигомерной организацией. Наиболее изученными гемоглобинами являются: гемоглобин взрослого человека - HbA1, имеющий структуру a2b2; фетальный гемоглобин HbF, состоящий из цепей a2g2; серповидноклеточный гемоглобин HbS, имеющий измененные b-цепи, вследствие чего его тетрамерная структура записывается как a2S2; минорный гемоглобин взрослого человека HbA2, представленный тетрамером a2d2. Четвертичная (олигомерная) структура наделяет гемоглобин важными особенностями, которые способствуют выполнению его уникальной биологической функции и обеспечивают возможность строгой регуляции его свойств. Гемоглобин связывает четыре молекулы кислорода на тетрамер (т.е. по одной на гем). Важнейшим его свойством является характерная кривая насыщения кислородом, которая имеет сигмоидную форму. Поэтому способность гемоглобина связывать О2 зависит от того, содержатся ли в данном тетрамере другие молекулы кислорода, т.е. последующие молекулы О2 присоединяются легче при частичной оксигенации гемопротеида. Это связано с конформационными перестройками в субъединицах тетрамера после оксигенирования одной из них. Присоединение О2 сопровождается разрывом солевых связей, образованных концевыми карбоксильными группами субъединиц. Указанные изменения заметно влияют на вторичную, третичную и особенно четвертичную структуру гемоглобина. В процессе оксигенации один ab-димер так поворачивается относительно другого ab-димера, что это приводит к компактизации тетрамера и повышению сродства гемов к кислороду. Кинетика кооперативного связывания кислорода важна не столько для взаимодействия О2 с гемоглобином, сколько для его максимально эффективной «отдачи» при тех парциальных давлениях О2, которые имеют место в периферических тканях.
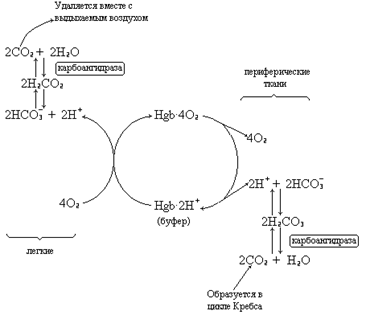
Гемоглобин не только переносит кислород от легких к периферическим тканям, но и ускоряет транспорт СО2 от тканей к легким. Данный гемопротеид связывает СО2 сразу же после высвобождения кислорода, при этом примерно 15% СО2, присутствующего в крови, преносится молекулами гемоглобина. Этот процесс зависит от присутствующей в эритроцитах карбоангидразы, которая катализирует превращение поступающего из тканей СО2 в угольную кислоту. Последняя быстро диссоциирует на бикарбонат-ион и протон.
Для предотвращения опасного понижения рН в крови должна существовать буферная система, способная поглощать избыток протонов. Эту функцию выполняетгемоглобин, который связывает два протона на каждые четыре освободившиеся молекулы кислорода и, тем самым, определяет буферную емкость крови. В легких идет обратный процесс: присоединение кислорода к дезоксигемоглобину сопровождается высвобождением протонов, которые связываются с бикарбонат-ионами с образованием угольной кислоты. Эффективно действующая карбоангидраза катализирует превращение угольной кислоты в углекислый газ, выдыхаемый из легких. Таким образом, связывание кислорода тесно сопряжено с выдыханием СО2. Этот обратимый процесс известен как эффект Бора (см. рис. 3.4). Эффект Бора является неотъемлемой частью общего механизма действия тетрамерного гемоглобина в качестве транспортного белка и определяется гем-гемовым взаимодействиями, лежащими в основе его кооперативных эффектов. Окисление ферро(Fe2+)-формы железа гема в ферри(Fe3+)-форму приводит к образованию метгемоглобина – функционально неактивной формы гемоглобина, лишенной возможности связывать кислород. Обычно в эритроцитах накапливается до 1% метгемоглобина в сутки, что связанно с обычным процессом автоокисления нормального гемоглобина. Восстановление гемового железа до ферро(Fe2+)-формы осуществляется NADH -метгемоглобинредуктазой, которая ответственна за поддержание низкого уровня физиологически неактивной формы гемоглобина в красных кровяных клетках. Вторым продуктом автоокисления гемоглобина являются высокоактивные супероксидные анионы – О2 · ¯, которые служат субстратом для супероксиддисмутазы, обеспечивающей прерывание цепи генерирования свободных радикалов в эритроцитах.
Рис. 3.4 Схема, отражающая механизм переноса углекислого газа гемоглобином (эффект Бора).
Выделение гемоглобина
Гемоглобин является наиболее доступным для лабораторного изучения гемопротеидом. Этот белок выделяют из эритроцитов после их лизиса при добавлении дистиллированной воды, эфира или толуола. В гипотонической среде эритроциты набухают, разрушаются и гемоглобин переходит в водный раствор. Эфир способствует растворению липидной оболочки эритроцитов, что облегчает выход их содержимого в раствор. Отделение белков плазмы достигается их осаждением полунасыщенным раствором сульфата аммония. После центрифугирования раствора и удаления осадка в супернатанте происходит медленная кристаллизация гемоглобина, которую осуществляют в течение длительного времени (около суток) на холоду.
|
||||||||||
|
|
Последнее изменение этой страницы: 2021-03-09; просмотров: 229; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.146.176.191 (0.012 с.) |

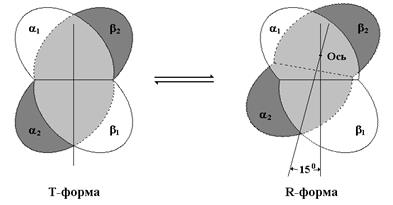 Четвертичная структура дезоксигемоглобина опи-сывается как Т-состояние, в то время как полностью оксигенированному гемо-глобину соответствует R-состояние. Переход гемо-глобина из Т- в R-форму сопровождается поворотом одной пары жестко связан-ных ab-субъединиц на 15° относительно другой ab-пары. Нужно отметить, что оксигенация гемоглобина сопровождается изменениями структуры в окружении гемогруппы. При оксигенировании атом железа, который в дезоксигемоглобине слегка выступает из плоскости гемового кольца, втягивается в плоскость гема. Вслед за атомом железа ближе к гему перемещается и проксимальный гистидин, а так же соседние аминокислотные остатки.
Четвертичная структура дезоксигемоглобина опи-сывается как Т-состояние, в то время как полностью оксигенированному гемо-глобину соответствует R-состояние. Переход гемо-глобина из Т- в R-форму сопровождается поворотом одной пары жестко связан-ных ab-субъединиц на 15° относительно другой ab-пары. Нужно отметить, что оксигенация гемоглобина сопровождается изменениями структуры в окружении гемогруппы. При оксигенировании атом железа, который в дезоксигемоглобине слегка выступает из плоскости гемового кольца, втягивается в плоскость гема. Вслед за атомом железа ближе к гему перемещается и проксимальный гистидин, а так же соседние аминокислотные остатки.