Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Развитие плодов, строение околоплодника. Разнообразие плодов, их классификация и эволюция. Гормональная регуляция созревания семян и околоплодника.Содержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте
Плоды характерны только для цветковых растений. Завязь - нижняя часть пестика, содержащая семяпочки, разрастается и превращается в плод. Таким образом, число семян, содержащихся в плоде, соответствует числу семяпочек. В строго ботаническом смысле плод-это зрелая завязь,содержащая семена-зрелые семяпочки.Стенка завязи преобразуется в околоплодник, а семязачатки-в семена. Околоплодник защищает семена от высыхания, механических повреждений, поедания и способствует распространению семян. Настоящий плод развивается только из завязи пестика. Плод, образующийся из чашелистиков, лепестков или цветоложа, называется ложным плодом. Так, плоды яблони состоят преимущественно из разросшегося мясистого цветоложа; только сердцевина яблока происходит из завязи. Настоящие и ложные плоды могут быть трех категорий: простые плоды (вишня, финик), образующиеся из цветка с одним пестиком; сложные плоды (малина, ежевика), развивающиеся из цветка с несколькими пестиками, и соплодия, образующиеся в результате срастания плодов одного или нескольких соцветий (ананас). Выделяют следующие виды плодов. 1). Сухие плоды: листовка - сухой, многосемянный (реже одно- или двусемянный), вскрывающийся в основном по брюшной стороне (купальница, пион); боб отличается от листовки тем, что раскрывается одновременно по брюшному шву и вдоль спинки (акация белая); стручок (стручочек) вскрывается снизу вверх с натянутой между створками перегородкой, на которой располагаются семена(сурепка, капуста);крылатка-по краям околоплодника образуется тонкая окраина в виде крыла (клен);семянка-нераскрывающийся плод(астра);зерновка-нераскрывающийся односемянный плод с тонким околоплодником, тесно прилегающим к кожуре(злаки);орешек-с кожистым или деревянистым околоплодником(земляника, гречиха, розы);орех-с деревянистым околоплодником, односемянный(лещина, грецкий орех);коробочка-многосемянный плод(табак). Сочные плоды: костянка-внутри деревянистая твердая косточка, за которой следует сочный мясистый слой, а снаружи плотный слой(слива, вишня, черешня, черемуха); ягода - сочный многосемянный плод (виноград, томаты, брусника, черника, клюква);яблоко-наружная часть околоплодника сочная, мясистая; внутренняя - перепончатая или хрящеватая (яблоня, груша, рябина, боярышник); тыквина - многосемянный плод с твердой кожурой, мясистым средним слоем и сочным внутренним (тыква, дыня);померанец - околоплодник толстокожистый, снаружи железистый, внутри волокнистый с жидким соком (апельсин, лимон).2). Вскрывающиеся и невскрывающиеся. 3). Односемянные и многосемянные. Сущ. классификация по типу генецея:1.простые(листовка,боб,орешек,костянка)2.сложные (многолистовка, многоорешек, многокостянка).Сухие:вскрывающиеся-коробочка,стручок,стручочек; невскрыв-ся-орех,жёлудь,зерновка,семя.Сочные:ягода,яблоко,помиранец,цитрусовые,тыквенные. Фитогормоны -это соед.,с помощью кот.-ых происход. взаимодействие кл-к, тканей и органов. Они.в.малых.кол-ах.необходимы.для.регуляции.Были.открыты:этилен,ауксин,цитокинин, гиберрилины, абсцезины.Они влияют на рост,созревания,старения,стрессы и транспорт вещ-в.Ауксины стимулируют корнеобр./обр-ие партенокарпных плодов,ускор-ют проростание семян.Ауксин.активирует деление и растяжение кл-к,необходим для формир-ия провод. пучков и корней, способствует разрастанию околоплодника. Гибберилины усиливают рост стеблей, обр. бессемянных плодов, с помощью их можно выводить растения из состояния покоя и ускор-ть процесс прорастания семян. Этилен участвует в процессе ускор-ет, созревания семян, также ускор. прорастание пыльцы семян. Абсцезиновая кис-та - это ингибитор роста. Цитакинины активируют рост клеток растяжения. Сохранение и распространение вида связаны с распространением семян и плодов. В процессе эволюции выработались различные приспособления: перенос семян и подов с помощью воды, ветра, животных или путем саморазбрасывания. 18. Рост и развитие растений. Гормоны растений как основные регуляторы роста и развития. Роль факторов внешней среды и гормонов в прохождении этапов развития. Каждый живой организм подвергается постоянным количественным и качественным изменениям, которые прекращаются только при известных условиях периодами покоя.Рост–необратимое увеличение размеров кл., органов, тканей, кот. сопровожд-ся новообразованием структур. Развитие – качественные изменения в растительном орг-ме, кот.сопровожд-ся заложением нов. органов, либо каких-то структур. Этапы развития растений: 1. зародышевый – вкл. в себя временной промежуток от момента образвания семени до созрев-я зародыша. 2.ювинеальный (молодость) – вкл. в себя период от прорастания семян и закан-ся формир-ем вегетат-х органов. 3.репродуктивный (зрелости) – нач-ся с момента закладки генеративных органов, вкл. в себя опыление, оплодотворение и развитие плодов и семян. 4.старение и смерть – хар-ся снижением жизненной активности, стеранием его органов и постепенное их отмирание. Покой растений – временное снижение функциональной активности растений. Даже у тропич-х растений все равно наблюд-ся покой. Бывает: вынужденным (опред-ся неблагопр-ми условиями внеш. среды) и физиологическим (глубокий, опред-ся внутр. факторами орг-ма). Для рестений, кот.ведут относительно неподвиж. образ жизни, но испыт-т повреждения-характерна регенерация, т.е. восстановление поврежденных уч-ков. М.б. физиологическая и травматическая. Факторы, влияющие на рост и развитие: Внутренние: возраст растений, развитость фотосинтетической ткани, наличие в раст. воды, интенсивность обменных процессов, интенсивность дых-я. Внешние – наличие пит-х элементов, воды в почве, температура, освещенность. Фитогормоны – это природные регуляторы роста, кот.оказывают влияние на морфогенетич. процессы растений. Делят на 2 группы:1 стимуляторы роста (ауксин, цитокенин, гиберелин).2.ингибиторы роста (этилен и абсцизовая кислота). Ауксин – синтезир-ся в верхушечных почках главных побегов, обеспеч-т апикальное доминирование(верхушечный рост),сдерживает рост боковых побегов, обеспеч. рост клеток растяжения, а также стимулир-т рост корней. Гормоны передвиг-ся в растении по живым элементам флоэмы, проникает ч/з оболочку кл., увлекает за собой протоны Н, кот.подкисляют оболочки кл., что повышает кислотность кл. стенки-активирует кислые гидролазы, кот. разрывают поперечные сшивки м/ду мол. целлюлозы, вода поступает в кл., оказ-ет давление на цит-му. Цитокинины– в 63г. получен из незрел.зерновок кукурузы – зеротин. стимулирует деление кл. Синтезир-ся в точке роста и рапростр-ся по растению в составе ксилемного тока.при увеличении концентрации цитокенинов увелич-ся синтез нуклеин-х кислот. При нач-х этапах старения органа обработка цитокенином приводит к омоложению. Гибберелин– регулирует рост. Его действие отлич-ся от действия ауксина. Выводит органы раст. из состояния покоя.Синтезир-ся в листьях, семенах, корнях.Передвиг-ся по проводящим тканям ксолемного ифлоэмного тока. Абсцизовая к-та– гормон абсцизин.Синтез-ся г.о. в листьях. Сдержиывавет рост, переводит растение в состояние покоя, снижает синтез АТФ, всех физиолог-х процессов, стимулирует закрытие устьиц. Этилен– в низких концентрациях обладает гормональным оздействием. Стимулирует созревание плодов, развитие отделительного слоя в черешках листьев и в местах крепления плодов. Содержание этилена регулир-ся самим растением. 19. Фотосинтез. Химизм процесса фотосинтеза. Возникновение и эволюция фотосинтеза. Фотоси́нтез — это процесс образования органического вещества из углекислого газа и воды на свету при участии фотосинтетических пигментов (хлорофилл у растений, бактероихлорофилл и бактериородопсин у бактерий). В современной физиологии растений под фотосинтезом чаще понимается фотоавтотрофная функция — совокупность процессов поглощения, превращения и использования энергии квантов света в различных эндэргонических реакциях, в том числе превращения углекислого газа в органические вещества. Общая упрощенная формула фотосинтеза выглядит так: 6СО2 +6Н2О+энергия света=С6Н12О6 +6О2↑ Различают оксигенный и аноксигенный типы фотосинтеза. Оксигенный гораздо более широко распространён, его осуществляют растения, цианобактерии и прохлорофиты. В данной статье описан только он, аноксигенному фотосинтезу пурпурных и зелёных бактерий, а также геликобактерий посвящена отдельная статья. Фотосинтез — сложный многоступенчатый процесс; реакции фотосинтеза подразделяют на две группы: реакции световой фазы и реакции темновой фазы. Световая фаза происходит только в присутствии света в мембранах тилакоидов при участии хлорофилла, белков-переносчиков электронов и фермента — АТФ-синтетазы. Под действием кванта света электроны хлорофилла возбуждаются, покидают молекулу и попадают на внешнюю сторону мембраны тилакоида, которая в итоге заряжается отрицательно. Окисленные молекулы хлорофилла восстанавливаются, отбирая электроны у воды, находящейся во внутритилакоидном пространстве. Это приводит к распаду или фотолизу воды. Темновая фаза протекает в строме хлоропласта. Для ее реакций не нужна энергия света, поэтому они происходят не только на свету, но и в темноте. Реакции темновой фазы представляют собой цепочку последовательных преобразований углекислого газа (поступает из воздуха), приводящую к образованию глюкозы и других органических веществ. Выделяют три этапа фотосинтеза: фотофизический, фотохимический и химический. На первом этапе происходит поглощение квантов света пигментами, их переход в возбуждённое состояние и передача энергии к другим молекулам фотосистемы. На втором этапе происходит разделение зарядов в реакционном центре, перенос электронов по фотосинтетической электронотранспортной цепи, что заканчивается синтезом АТФ и НАДФН. Первые два этапа вместе называют светозависимой стадией фотосинтеза. Третий этап происходит уже без обязательного участия света и включает в себя биохимические реакции синтеза органических веществ с использованием энергии, накопленной на светозависимой стадии. Чаще всего в качестве таких реакций рассматривается цикл Кальвина и глюконеогенез, образование сахаров и крахмала из углекислого газа воздуха. Фотосинтез присущ всем зеленым растениям, циано- и некоторым другим бактериям. Роль фотосинтезирующих организмов в природе определяющая: будучи «производителями» органических веществ из неорганических, они обеспечивают все другие организмы «строительным материалом» и энергией, а также необходимым для дыхания кислородом. Согласно общепризнанной в настоящее время эволюционной теории происхождения и развития жизни, которая более 50 лет назад была сформулирована А. И. Опариным, первичные, способные к самовоспроизводству живые образования возникли в результате абиогенной химической эволюции. Будучи окруженными близкими по составу, но еще неживыми органическими соединениями, эти первичные существа могли осуществлять в бескислородной среде анаэробный гетеротрофный тип питания с помощью небольшого набора ферментов. Постепенное истощение и деградация органических веществ, синтезированных абиогенным путем, сопровождались накоплением все более окисленных соединений, вплоть до появления наиболее бедного энергией соединения углерода — углекислоты. Это влекло за собой необходимость все большего и большего совершенствования и усложнения ферментативного аппарата, необходимого для ассимиляции все более окисленных веществ. В этих условиях, которые все еще характеризовались отсутствием в среде кислорода, вполне вероятно возникновение первичных автотрофных организмов, которые осуществляли восстановление углекислоты за счет химической энергии, полученной из минеральных веществ. Такой тип питания получил название хеморедукции. Среди современных организмов известна группа сульфатредуцирующих микроорганизмов, которые восстанавливают сульфаты до сероводорода, используя для этой цели молекулярный водород. Появление в этот период, который характеризовался сильно восстановительными условиями среды, светпоглощающих пигментов фотосенсибилизаторов привело, очевидно, к замене химической энергии в процессах хеморедукции на световую. Возник простейший тип фотоавтотрофного питания, который получил названи фоторедукции и бактериального фотосинтеза. Такой тип питания осуществляют современные фототрофные бактерии— пурпурные серобактерии (Thiorhodaceae) и зеленые серобактерии (Chlorobacteriaceae), у которых роль пигмента-фотосенсибилизатора выполняет бактериохлорофилл и которые являются строгими анаэробами. Пурпурные и зеленые серобактерии восстанавливают углекислоту за счет энергии света, используя в качестве Н-донора сероводород (H2S):
Представленное итоговое уравнение бактериального фотосинтеза (фоторедукции) очень напоминает, как мы видим, приведенное выше суммарное уравнение фотосинтеза хлорофиллоносных растений. В результате сравнительного анализа Ван-Ниль показал, что оба эти процесса могут быть записаны в общем виде одним итоговым уравнением:
где Н2А — донор водорода, в качестве которого фотосинтезирующие бактерии используют сероводород, а остальные растения — воду. Вода является более окисленным соединением по сравнению с сероводородом. Использование ее в качестве донора водорода связано с необходимостью дополнительной затраты энергии и стало возможно благодаря дальнейшему совершенствованию фотохимического аппарата, которое состояло в появлении у растений (начиная с сине-зеленых водорослей) хлорофилла (вместо бактериохлорофилла) и дополнительной фотохимической системы, так называемой «фотосистемы П». Использование воды в качестве донора водорода привело к тому, что в процессе фотосинтеза стал выделяться кислород, что, в свою очередь, ознаменовало переход от анаэробной к аэробной жизни на нашей планете. На эволюционную связь фото редукции и фотосинтеза может указывать способность ряда сине-зеленых, зеленых, красных и бурых водорослей обратимо переходить к фоторедукции при переводе их в анаэробные условия в атмосферу водорода. Таким образом, фотоавтотрофный тип питания и фотосинтез возникли в процессе эволюции как «надстройка» над первичным гетеротрофным типом питания. Появление на Земле фотосинтеза было обусловлено всем ходом предшествовавшей биологической эволюции и явилось поворотным пунктом в переходе от анаэробного к аэробному типу обмена веществ. Рассмотренная схема дает представление лишь об общих чертах эволюции фотосинтеза и является в значительной степени гипотетической. Многие этапы эволюции фотосинтеза и тем более ее детали остаются неясными, ряд моментов по-разному интерпретируется учеными.
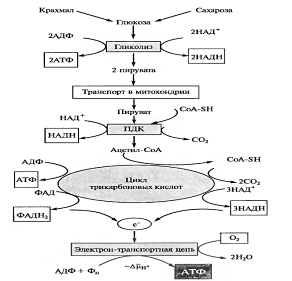
20. Дыхание как энергетический процесс, его механизмы и значение в жизни растительного организма. Пути дыхательного обмена и эволюция дыхания. Дыхание — один из важнейших процессов обмена веществ растительного организма. Выделяющаяся при дыхании энергия тратится как на процессы роста, так и на поддержание в активном состоянии уже закончивших рост органов растения. Вместе с тем значение дыхания не ограничивается тем, что это процесс, поставляющий энергию. Дыхание, подобно фотосинтезу, сложный окислительно- восстановительный процесс, идущий через ряд этапов. На его промежуточных стадиях образуются органические соединения, которые затем используются в различных метаболических реакциях. К промежуточным соединениям относят органические кислоты и пентозы, образующиеся при разных путях дыхательного распада. Таким образом, процесс дыхания — источник многих метаболитов. Несмотря на то, что процесс дыхания в суммарном виде противоположен фотосинтезу, в некоторых случаях они могут дополнять друг друга. Оба процесса являются поставщиками как энергетических эквивалентов (АТФ, НАДФН), так и метаболитов. Как видно из суммарного уравнения, в процессе дыхания образуется также вода. Эта вода в крайних условиях обезвоживания может использоваться растением и предохранить его от гибели. В некоторых случаях, когда энергия дыхания выделяется в виде тепла, дыхание ведет к бесполезной потере сухого вещества. В этой связи при рассмотрении процесса дыхания надо помнить, что не всегда усиление процесса дыхания является полезным для растительного организма. Значение дыхания в жизни растения. Дыхание — один из центральных процессов обмена веществ растительного организма. Значение дыхания не ограничивается тем, что это процесс, поставляющий энергию. Дыхание, подобно фотосинтезу, сложный окислительно-восстановительный процесс, идущий через ряд этапов. На его промежуточных стадиях образуются органические соединения (органические кислоты и пентозы), которые затем используются в различных метаболических реакциях. Т. о., процесс дыхания — важнейший источник многих метаболитов. Как видно из суммарного уравнения, в процессе дыхания образуется вода. Исследования показали, что эта вода в крайних условиях обезвоживания может быть использована растением и предохранить его от гибели. Благодаря всем этим особенностям дыхание — это центральный метаболический процесс, переплетающийся многочисленными связями с другими процессами обмена. Процесс дыхания противоположен фотосинтезу. Если фотосинтез — синтетический процесс образования органического вещества, то дыхание — процесс распада, т. е. траты органического вещества. В некоторых случаях, когда энергия дыхания выделяется в виде тепла, дыхание ведет к бесполезной потере сухого вещества. Не всегда усиление процесса дыхания является полезным для растительного организма. Теории Дыхания. 1 теория Баха – суть: молекулярный кислород имеет двойную связь и для того чтобы его активировать, необходимо эту двойную связь расщепить. Легко окисляющееся соединение А взаимодействует с кислородом и, разрывая двойную связь, образует пероксид АО2. Т. о., по мысли Баха, активация кислорода есть образование пероксида. В свою очередь пероксидное соединение, взаимодействуя с соединением В, окисляет его; затем эта реакция повторяется со вторым атомом кислорода и второй молекулой соединения В. Получается полностью восстановленное исходное соединение — акцептор кислорода А и полностью окисленное вещество В: 2 теория Палладина. Он считал, что О2 не является непосредственным окислителем субстрата, а всего лишь конечный акцептор е —, который является замыкающим звеном в ЭТЦ дыхания. Палладин предположил, что в процессе дыхания используется дыхательные пигменты – это те же вещества восстанавливаясь могут изменять свой цвет. Общая схема процесса дыхания (основное уравнение дыхания на анаэробную (1) и аэробную (2) части): 1) С6Н12О6 + 6Н2О + 12R = 6СО2 + 12R Н2; 2) 12R Н2 + 6О2 = 12R + 12Н2О С6Н12О6 + 6О2 = 6СО2 + 6Н2О где R — это окрашенный дыхательный пигмент, способный отнимать водород от субстрата, a RH2 — бесцветный дыхательный хромоген. Существуют две основные системы и два основных пути превращения дыхательного субстрата, или окисления углеводов: 1) гликолиз + цикл Кребса (гликолитический); дихотомический
2) пентозофосфатный (апотомический). Данный путь дыхательного обмена является наиболее распространенным и, в свою очередь, состоит из двух фаз. Первая фаза – анаэробная (гликолиз), локализована в цитоплазме. Вторая фаза – аэробная, локализована в митохондриях. В процессе гликолиза происходит преобразование молекулы гексозы до двух молекул пировиноградной кислоты (ПВК): С6Н12О6 → 2 С3Н4О3 + 2Н2 Вторая фаза дыхания – аэробная - требует присутствия кислорода. В эту фазу вступает пировиноградная кислота. Общее уравнение этого процесса можно представить так: 2ПВК + 5 О2 + Н2О → 6СО2 + 5Н2О Относительная роль этих путей дыхания может меняться в зависимости от типа растений, возраста, фазы развития, а также в зависимости от факторов среды. Процесс дыхания растений осуществляется во всех внешних условиях, при которых возможна жизнь. Растительный организм не имеет приспособлений к регуляции температуры, поэтому процесс дыхания осуществляется при температуре от -50 до +50°С. Нет приспособлений у растений и к поддержанию равномерного распределения кислорода по всем тканям. Именно необходимость осуществления процесса дыхания в разнообразных условиях привела к выработке в процессе эволюции разнообразных путей дыхательного обмена и к еще большему разнообразию ферментных систем, осуществляющих отдельные этапы дыхания. При этом важно отметить взаимосвязь всех процессов обмена в организме. Изменение пути дыхательного обмена приводит к глубоким изменениям во всем метаболизме растений. 21. Водоросли как экологическая группа низших растений. Особенности строения, питания и размножения. Основные отделы, роль в эволюции растительного мира. В. - большая группа низших растений, в подавляющем большинстве обитающих в воде, где они составляют основную массу растительности. Некоторые прикрепляются ко дну водоемов или подводным предметам и составляют фитобентос, большинство же находится в толще воды во взвешенном состоянии или плавает - они составляют фитопланктон. Часть водорослей живет в почве, на почве или на стволах деревьев. Способы питания:- фототрофный –происходит благодаря процессу фотос-за, возможно только при условии наличии пигмента акцептора света (хромотофора),- гетеротрофный (готовыми орг-ми в-вами), присущ некот. с-з, желтыми диатомовым, - миксотрофный (смешанный), в зависимости от условий питания (эвглена зел.).Типы организации талломов: амебоидный, одноклеточный, кокоидный (коллониальный), нитчатый, гетеротрихиальный, пластинчатый, сифональный (одкон. особь, но многоядерная), харофитный (различают узлы и междуузлия). Размножение –способность орг-мов воспроизводить себе подобных. 3 способа:- вегетативное (у 1-кл. происходит деление кл. пополам, у многокл. путем деления особи на отдельные части, путем образования специализир. стр-р – выводковые почки, крахмалоносные клубеньки), - бесполое –деление протопласта на части, их обособление и выход во внеш. среду. Обособленные части протопласта образуют споры.У многих В. споры имеют жгутики–зооспоры, кот. образ-ся в зооспорангиях.После выхода из спорангия они активно плавают, теряют жгутики, одев-ся плотной обол-кой и дают начало нов. особи. - половое – слияние 2-х половых кл. (гамет) с образованием зиготы (2n). Несколько типов пол.процесса:1. хологамия – слив-ся неспециализир-е кл. 2-х однотипных особей (у бесжгутиковых наз-ся конъюгацией - зигогамией). 2. изогамия – обе одинаковые и обе подвижны. 3. гетерогамия – обе подвижные, с одинаковым набором хромосом. 4. оогамия – слив-ся женская, более крупная, неподвижная с более мелкой, подвижной. Споры и гаметы могут развиваться в клетках одной и той же особи, но возможно развитие спор на одной, а гамет - на другой. В таком случае особь, на которой развиваются споры, называется спорофитом, а особь, на которой образуются гаметы, - гаметофитом. При слиянии гамет образ-ся зигота, кот. в большинстве случаев превращ-ся в покоящуюся спору, кот. спустя некот. время прорастает и дает нач. нов. особи. Известно более 40 тыс. видов водорослей, которые объединяются в два полцарства — Багрянки и Настоящие водоросли. Последние подразделяются на несколько обособленных отделов (Зеленые, Харовые, Диатомовые, Золотистые, Бурые и др.), которые отличаются друг от друга по ряду таких важнейших признаков, как структура таллома, набор фотосинтезирующих пигментов и запасных питательных веществ, особенности размножения и циклы развития, местообитание и т. п. Водорослям присуще исключительное морфологическое разнообразие. Они бывают одноклеточные (хламидомонада, хлорелла), колониальные (вольвокс) и многоклеточные, среди которых встречаются нитчатые (спирогира, улотрикс) и пластинчатые (ульва, ламинария и др.). Каждая из этих форм имеет различные размеры — от I мкм до нескольких десятков метров (некоторые бурые и красные). Основные отделы водорослей: зеленые, бурые, красные. Зеленые: сам.крупный отд. из водорослей от 13 до 20 тыс. видов. по окраске чисто зелен. цвета (хромотофор). Хр-ры различной формы, чаще крупные.Пигменты хлорофилл А,В, альфа- бета- каротины и ксантофиллы (более 10). имеют все типы организации таллома, кроме амебоидного. Кл. 1-ядерные, реже многояд-ные. В состав кл. оболочки входят целлюлоза и пектиновые в-ва. Размножение: вегетативное, у 1-клет-х происходит путем деления кл. пополам, у колониальных–распада на отдельные уч-ки, у многоклет-х– частью таллома, у нитчатых–путем деления на отдельные нити, у пластинч-х–образованием специализ-х стр-р–выводковых почек. Бесполое–посредством зооспор, кол-во жгутиков кратно 4. Половое–холо-, изо-, оо-, гетерогамии, конъюгации (зигогомия). гаметы с 2, реже с 2 жгутиками.Зигота у большинства одев-ся толстой оболочкой–циста и прорастает после длительного периода покоя. Стадия покоя–ЗИГОТА. Большинство живет в пресных водоемах, др. на скалах, деревьях, вступает в симбиоз с грибами=лишайники. Значение –корм для фауны, уч-ют в круговороте в-в, употребл-ся в пищу. Представители: хламидомонада, вольвокс, улотрикс, космариум, спирогира и др. Бурые: в основном морские. Преобладают на каменистых побережьях, в холодных районах земного шара. Размеры от неск. мм до неск. десятков м. По внеш. виду м.б. ветвистыми, кустистыми, корочковыми, пластинчатыми, в виде шнуров, лент, сложно рассеченных на стеблевую и листовую часть. Слоевище некот. крупн. представителей имеют специализир. воздушные вздутия для поддержания талломов в толще воды. У них есть ассимиляционная, запасающая, проводящая, механическая ткани. Нет 1-клет-х, колониальных и 1-нитчатых форм организации таллома. Оболочки кл. ослизнены, состоят из пектонивых в-в и гемицеллюлозы.Хромотофоры содержат пигменты: хлорофилл А,С, каротины, фукоксантин. Продукты запаса–углевод ламинорин, масла и 6-атомный спирт манит. Размн-ие вегетативное (уч-ми таллома и специализ. выводковыми почками), бесполое (зооспоры и апланоспор).Половое (изо-, гетеро-, оогамия). у всех бурых яйцеклетка оплодотвор-ся вне оогония, за счет хемотаксиса. Зигота без периода покоя прорастает в спорофит. Значение –пища, место размножения и укрытия многих видов жив-х, субстратом для макро- и микроорг-мов, орган-х удобрений. В пишевой, парфюмерной, текстильной промышленности. Из них получают агар-агар, спирт манит. Красные: Распространены в морях, некоторые виды встречаются в пресных водах и на почве. Строение таллома красных сходно со строением талломов бурых водорослей. Их окраска обусловлена такими пигментами, как хлорофилл, фикоэритрин, фикоциан (получили из за них свое название).Хроматофоры у красных водорослей имеют вид дисков, пиреноидов нет. Запасные продукты содержатся в них в виде масла и специфического багрянкового крахмала. Таллом имеет вид кустиков, составленных из многоклеточных ветвящихся нитей, реже пластинчатый или листовидный до 2 м длины. Пектиново-целлюлозные клеточные стенки у некоторых видов ослизняются настолько сильно, что весь таллом приобретает слизистую консистенцию. Поэтому некоторые виды применяются для получения агар-агара,широко используемого в пищевой промышленности. Клеточные стенки некоторых красных водорослей могут быть инкрустированы углекислым кальцием и углекислым магнием, что придает им твердость камня. Такие водоросли участвуют в образовании коралловых рифов. В странах Восточной Азии некоторые виды порфир употребляют в пищу. В Норвегии во время морского отлива на прибрежную часть, богатую красными водорослями, выпускают овец, как на пастбище. 22. Отличительные признаки моховидных, классификация, роль в природе и хозяйственное использование. Мхи насчит-т около 25 тыс. видов. Относ-ся к высшим споровым раст., для кот. хар-но разделение тела на органы. Встреч-ся повсеместно, но преимущественно в обл. с умерен. и хол. климатом: в густых лесах, на болотах, на стенах и крышах, на скалах. ОБЩАЯ ХАР-КА ОТДЕЛА: 1. чередование поколений с преобладанием в цикле развития гаметофита. 2. у мхов нет настоящих проводящих элементов, только предшественники про-х эл-тов: гидроиды (проводят воду) и липтоиды (пров. орг-кие в-ва).3.вегет-ное тела представлено слоевищем. Настоящих корней и листьев у мхов нет, имеются ризоиды.4.спорофит прикрепл-ся к гаметофиту, полностью зависит от него и пит-ся за счет него.5.споры образ-ся в споровой коробочке – спорогон. 6.оплодотворение происходит при наличии воды. Значение. Участвуют в образовании лесных фитоценозов, способны накапливать вредные в-ва из почвы. Явл-ся регулятором водного баланса. Сфагнум уч-ет в торфообразовании, обладает антисептическими св-ми и препятствует развитию гнилостных бактерий (в-во сфагноль), способствует мумизации. Мохообразные обычно входят в состав первичных растительных группировок на поверхно-сти скал и камней, они часто являются пионерами зарастания углублений, заполненных водой, и обнаженных почв. Постепенно отмирая, пионерные виды мохообразных подготавливают субстрат для поселения других видов мохообразных или сосудистых растений. Иногда роль мохообразных в сообществе сильно возрастает в связи с резким изме-нением условий местообитания, вызванным воздействием животных, человека и факторов, обусловленных жизнедеятельностью самого сообщества растений. Например, уничтожение леса этого мощного насоса, выкачивающего влагу из почвы, в бессточных местах или в мес-тах со слабым стоком и близким к поверхности уровнем грунтовых вод приводит к появле-нию в почве избыточной влаги. Начинается процесс заболачивания. На почве, покрытой до вырубки, например, черникой, лесными травами и некоторыми лесными мхами, появляется сплошной покров из мха политриха обыкновенного Polytrichum commune. Затем появляются сфагновые, роль которых в растительном покрове постепенно возрастает, и наконец образуется моховое болото. Классификация. Отдел моховидные делится на 3 класса: антоцеротовые, печеночные и листостебельные мхи. Класс печеночные мхи – Hepaticopsida. Тело представлено слоевищем в форме пластинки дорзовентрального строения. От нижней (вентральной) стороны отходят одноклеточные ризоиды, соединяющие растение с субстратом. Представителем может служить маршанция. Слоевище ее покрыто эпидермой. Клетки нижней стороны образуют выросты - ризоиды. Верхняя эпидерма имеет дыхательные отверстия - дыхальца. К верхней эпидерме примыкает ассимиляционная ткань с крупными воздушными полостями, к нижней - запасающая ткань. Растение разнополое. На женских экземплярах развиваются женские подставки с архегониями, на мужских -мужские подставки с антеридиями. В старину маршанция - Marchantia polymorpha использовалась для лечения болезней печени. Анатомическое стр-е слоевища: слоевище покрыто однослойной эпидермой. В верхней эпидерме располаг-ся устьица, кот постоянно открыты, дальше располаг-ся основная ткань, образованная паренхимными кл. Снизу также покрыто 1-слойной эпидермой, в кот. имеется 2 типа ризоидов.. Размножение: вегетативное. На слоевище с верх-й стороны образ-ся выводковые корзинки, в кот. формир-ся выводковые почки, кот. при созревании вымываются с током воды. Прикрепл-ся к субстрату и дают начало нов. талому. Половое – цикл развития – различают муж. и жен. гаметофиты. На жен. формир-ся жен. Подставки, на муж. муж. подставки. На лопастях жен. подставок с нижней стороны формир-ся архегонии с яйцекл., на муж. с верхней стороны – антеридии со спермат-ми. При наличии воды = оплодотворение = зигота = спорогон (спороносная коробочка). Класс Листостебельные мхи (BRYOPSIDA, ИЛИ MUSCI) Листостебельные мхи – самый крупный класс моховидных. Гаметофит листостебельных мхов расчленен на стебель и листья. Стебель имеет радиальное строение, на нем тесной спиралью располагаются листья. На стебле формируются органы полового размножения – антеридии и архегонии. Спорофит (спорогон) развивается из оплодотворенной яйцеклетки и состоит из коробочки, ножки и гаустории (стопы). В коробочке формируются только споры, элатеры отсутствуют. Со споры развивается протонема (предросток) в виде длинной разветвленной нити, реже она пластинчатая. Представитель - Кукушкин лен. Состоят из стебля, на кот. по спирали располаг-ся чешуйчатые, редуцированные листья. Внутри стебля гидроиды, снаружи – липтоиды. Прикрепл-ся ризоидами. Лист с нижней стороны – 1-слойная эпидерма, далее склеренхима (мех-я ткань), затем проводящие эл-ты флоэмы и ксилемы. Все остальное пространство заполнено паренхимой, далее слой выстилающих кл., от кот. идут кл. ассимиляторы (ф-я фотосинтеза), кажд. Столбик заканч-ся покровной кл. Размножение - половое – различ-т муж. и жен. гаметофит. На верхушке жен. гаметофита формир-ся архегонии с яйцекл., на верхушке муж. – антеридии со сперматозоидами. При наличии воды = оплод-е = зигота (2n) = спорогон (коробочка), кот. состоит из центральной колонки по сторонам кот. формир-ся спорогонная ткань. Сверху колонка имеет расширение и образует перистом с зубчиками. Сверху коробочка прикрыта крышечкой. Для коробочки хар-рен фотосинтез, поэтому питание самостоятельное и прикрепл-ся к гаметофиту при помощи истинной ножки. В спорогенной ткани происходит мейоз = споры (n), кот прорастают в нитчатую протонему, на кот. формир-ся молодой гаметофит. Класс подразделяют на три подкласса: сфагновые (Sphagnidae), андреевые (Andreaeidae), бриевые (Bryidae). Класс Антоцеротовые (ANTHOCEROTOPSIDA) Свое название получили от греческих слов anthos – цветок, keros – рог. Спорогоны имеют вытянутую изогнутую в виде рога форму. Это талломные растения в виде розеткоподобной пластинки.Чертами примитивной организации антоцеротовых являются пластинчатые хлоропласты с пиреноидами, как у водорослей.
|
||||
|
|
Последнее изменение этой страницы: 2020-11-11; просмотров: 459; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.141.47.139 (0.019 с.) |

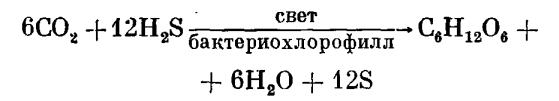 фотосинтеза
фотосинтеза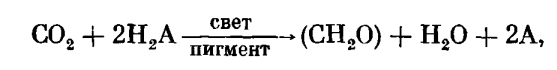 фотосинтеза
фотосинтеза