Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Взаимоотношения давлений в дыхательной системеСодержание книги
Поиск на нашем сайте
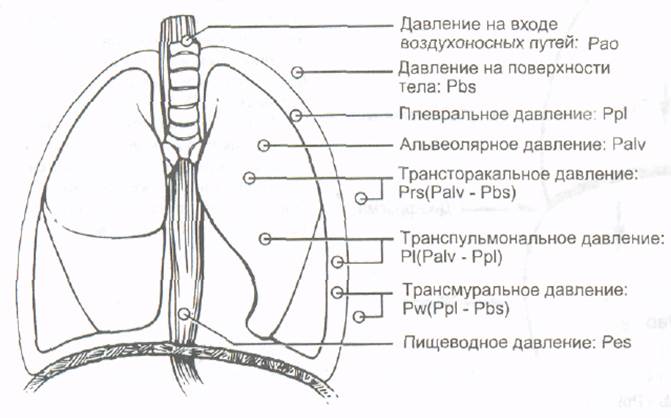
Различные виды давлений, определяющие поток воздуха в легких, схематически изображены на рис.13. Рао - давление на входе в воздухоносные пути (ВП) (т. е. в ротовой полости). В нормальных условиях, когда поток отсутствует (в конце вдоха и выдоха) и ВП открыты в атмосферу, Рао равно нулю. Pbs - давление атмосферы на поверхность тела. Некоторые механические системы для принудительной вентиляции легких создают отрицательное давление на поверхность тела, генерируя вакуум вокруг туловища пациента. В этих условиях Pbs прерывисто опускается ниже нуля. Ppl - плевральное давление - давление внутри плевральной полости. Его величина зависит от величин и направлений сил, создаваемых эластической паренхимой легких и грудной стенкой. Ppl может быть измерено с помощью баллонного катетера, помещенного внутрь пищевода, поскольку изменения внутрипищеводного давления во время дыхания отражают изменения внутриплеврального давления. Давление, создаваемое эластической паренхимой легкого, на рис. не представлено; оно направлено внутрь и называется давлением эластической отдачи, Pel. Альвеолярное давление, Palv - давление внутри альвеол. Оно может быть отрицательным (во время вдоха), положительным (во время выдоха) или нулевым (в конце вдоха и в конце выдоха, когда поток отсутствует и голосовая щель открыта). Альвеолярное давление представляет собой сумму давления эластической отдачи и плеврального давления: Palv=Pel + Ppl Pl - сквозное чрезлегочное или транспульмональное давление необходимо для возникновения воздушного потока и поддержания данного уровня расправления легких. Рl составляет разницу между альвеолярным и плевральным давлениями (Pl = Palv - Ppl). Pw- сквозное чрезстенное или трансмуралъное давление является разницей между плевральным давлением и давлением на поверхности тела (Pw = Ppl - Pbs). Prs - трансторакальное давление представляет разницу между альвеолярным давлением и давлением на поверхности тела (Prs = Palv - Pbs).
Рис.13. Схематическое изображение давлений и градиентов давлений, создающих поток воздуха. Регуляция дыхания. Дыхательный центр. Межреберные мышцы получают двигательную иннервацию от торакальной части спинного мозга, диафрагма иннервируется из его шейного отдела. Однако клеточные тела двигательных нейронов, расположенные в передних рогах спинного мозга, не образуют в своей совокупности дыхательного центра.
Исследования Н. А. Миславского показали, что с дыхательными движениями непосредственно связана область продолговатого мозга. Проводящие пути от этого центра спускаются по передним и переднебоковым столбам белого вещества спинного мозга и заканчиваются в его сером веществе на различных уровнях, соответствующих моторным клеткам нервов, идущих к диафрагме и межреберным мышцам. Импульсы, связанные с вдохом и выдохом, возникают в продолговатом мозгу и затем распространяются к подчиненным двигательным центрам спинного мозга или же, в случае вспомогательных дыхательных движений мышц носа и гортани, к двигательным центрам nn. vagi и facialis. Локализация дыхательного центра, описанная Миславским, полностью подтверждается современными исследованиями. Подтверждается также и его предположение о наличии двух отделов дыхательного центра: инспираторного и экспираторного. Центр является двусторонним, инспираторный отдел дыхательного центра занимает вентральную часть ретикулярной формации, экспираторный отдел - ее дорзальную часть, причем области центров вдоха и выдоха частично перекрывают друг друга. При действии электрическим током на центр инспирации ритмическое дыхание прекращается, так как возникает максимальный вдох, который длится непрерывно весь период раздражения. При раздражении экспираторного центра возникает такой же непрерывный выдох. Весьма существенно, что возбуждение экспираторного центра обусловливает торможение в центре инспираторном. Максимальный вдох, вызванный раздражением этого центра, подавляется одновременным раздражением центра экспирации, если оно достигает достаточной силы. Локализация дыхательного центра устанавливается различными методами, из них наиболее детальные сведения получены при прямом раздражении очень тонкими электродами вещества продолговатого мозга. При соблюдении определенных условий распространение тока при таком раздражении не превышает круга с радиусом 0,5 мм. Соответственно этому границы дыхательного центра и его инспираторного, и экспираторного отделов можно считать установленными с точностью до 0,5 мм.
Инспираторный отдел посылает импульсы к мотонейронам спинного мозга, составляющим ядрам диафрагмального и наружных межреберных нервов, в результате сокращаются мышцы вдоха. При возбуждении экспираторного центра прекращается генерация потенциалов действия в инспираторных нейронах и вдыхательные мышцы расслабляются. Дыхательный центр может возбуждаться автоматически, что обусловлено протекающими в нем процессами обмена веществ, а также высокой чувствительностью к углекислоте. Между центрами вдоха и выдоха устанавливаются нормальные отношения благодаря центру пневмотаксиса, расположенному в области варолиева моста, который осуществляет плавную ритмическую смену фаз дыхания, регулирует частоту и глубину дыхания. В регуляции дыхания принимают участие гипоталамус, а также кора больших полушарий, которые обеспечивают всю гамму тончайших приспособлений дыхания к потребностям организма. Дыхательные рефлексы. Раздражение афферентных нервов может обусловить учащение и усиление дыхательных движений или же, наоборот, замедление, или даже временную полную остановку дыхания. Дыхательный центр, следовательно, подвергается и рефлекторной стимуляции, и рефлекторному торможению. И тормозное, и стимулирующее воздействие могут быть получены с кожной поверхности тела (например, при погружении в холодную воду) под влиянием зрительных и слуховых раздражений, при эмоциональных реакциях организма, в случае прямого раздражения электрическим током среднего мозга и различных отделов коры больших полушарий. Значение кортикального контроля, обеспечивающего возможность произвольного управления дыхательными мышцами, делается особенно понятным, если вспомнить, что деятельность их служит не только для газообмена. Мышцы эти принимают участие в важнейшем для человека акте речи, связанном с прохождением воздушного потока через голосовую щель (звукообразование). Координационные процессы в центральной нервной системе, лежащие в основе образования звуков, представляют собой большую сложность, и в цепи этих процессов определенное место занимает управление деятельностью дыхательных мышц. Деятельность дыхательного центра может быть видоизменена рефлексом с любого чувствительного спинномозгового или черепномозгового нерва, а также под влиянием интрацентральных проводящих путей, идущих от высших отделов головного мозга. Реакции дыхательного центра могут быть разделены на группу защитных рефлексов и на рефлексы, участвующие в координации дыхательного акта. Рефлекторная регуляция дыхания осуществляется благодаря тому, что нейроны дыхательного центра имеют связи с многочисленными механорецепторами дыхательных путей и альвеол легких и рецепторов сосудистых рефлексогенных зон. В легких человека находятся следующие типы механорецепторов: 1) ирритантные, или быстроадаптирующиеся, рецепторы слизистой оболочки дыхательных путей; 2) рецепторы растяжения гладких мышц дыхательных путей; 3) J-рецепторы. Рефлексы со слизистой оболочки полости носа. Раздражение ирритантных рецепторов слизистой оболочки полости носа, например табачным дымом, инертными частицами пыли, газообразными веществами, водой вызывает сужение бронхов, голосовой щели, брадикардию, снижение сердечного выброса, сужение просвета сосудов кожи и мышц. Защитный рефлекс проявляется у новорожденных при кратковременном погружении в воду. У них возникает остановка дыхания, препятствующая проникновению воды в верхние дыхательные пути.
Рефлексы с глотки. Механическое раздражение рецепторов слизистой оболочки задней части полости носа вызывает сильнейшее сокращение диафрагмы, наружных межреберных мышц, а следовательно, вдох, который открывает дыхательный путь через носовые ходы (аспирационный рефлекс). Этот рефлекс выражен у новорожденных. Рефлексы с гортани и трахеи. Многочисленные нервные окончания расположены между эпителиальными клетками слизистой оболочки гортани и главных бронхов. Эти рецепторы раздражаются вдыхаемыми частицами, раздражающими газами, бронхиальным секретом, инородными телами. Все это вызывает кашлевой рефлекс, проявляющийся в резком выдохе на фоне сужения гортани и сокращение гладких мышц бронхов, которое сохраняется долгое время после рефлекса. Кашлевой рефлекс является основным легочным рефлексом блуждающего нерва. Рефлексы с рецепторов бронхиол. Многочисленные миелинизи-рованные рецепторы находятся в эпителии внутрилегочных бронхов и бронхиол. Раздражение этих рецепторов вызывает гиперпноэ (усиление дыхания), сужение бронхов, сокращение гортани, гиперсекрецию слизи, но никогда не сопровождается кашлем. Рецепторы наиболее чувствительны к трем типам раздражителей: 1) табачному дыму, многочисленным инертным и раздражающим химическим веществам; 2) повреждению и механическому растяжению дыхательных путей при глубоком дыхании, а также пневмотораксе, ателектазах, действии бронхоконстрикторов; 3) легочной эмболии, легочной капиллярной гипертензии и к легочным анафилактическим феноменам. Рефлексы с J-рецепторов. В альвеолярных перегородках в контакте с капиллярами находятся особые J-рецепторы. Эти рецепторы особенно чувствительны к интерстициальному отеку, легочной венозной гипертензии, микроэмболии, раздражающим газам и ингаляционным наркотическим веществам, фенилдигуаниду (при внутривенном введении этого вещества). Стимуляция J-рецепторов вызывает вначале апноэ (временную остановку дыхания), затем поверхностное тахипноэ (учащение дыхания), гипотензию и брадикардию. Рефлекс Геринга - Брейера. Раздувание легких у наркотизированного животного рефлекторно тормозит вдох и вызывает выдох. Перерезка блуждающих нервов устраняет рефлекс. Нервные окончания, расположенные в бронхиальных мышцах, играют роль рецепторов растяжения легких. Их относят к медленно адаптирующимся рецепторам растяжения легких, которые иннервируются миелинизированными волокнами блуждающего нерва. Рефлекс Геринга - Брейера контролирует глубину и частоту дыхания. У человека он имеет физиологическое значение при дыхательных объемах свыше 1 л (например, при физической нагрузке). У бодрствующего взрослого человека кратковременная двусторонняя блокада блуждающих нервов с помощью местной анестезии не влияет ни на глубину, ни на частоту дыхания. У новорожденных рефлекс Геринга - Брейера четко проявляется только в первые 3-4 дня после рождения.
Проприоцептивный контроль дыхания. Рецепторы суставов грудной клетки посылают импульсы в кору больших полушарий и являются единственным источником информации о движениях грудной клетки и дыхательных объемах. Межреберные мышцы, в меньшей степени диафрагма, содержат большое количество мышечных веретен. Активность этих рецепторов проявляется при пассивном растяжении мышц, изометрическом сокращении и изолированном сокращении интрафузальных мышечных волокон. Рецепторы посылают сигналы в соответствующие сегменты спинного мозга. Недостаточное укорочение инспираторных или экспираторных мышц усиливает импульсацию от мышечных веретен, которые через γ -мотонейроны повышают активность α-мотонейронов и дозируют таким образом мышечное усилие. Хеморефлексы дыхания. Напряжение кислорода и углекислого газа в артериальной крови человека и животных поддерживается на достаточно стабильном уровне, несмотря на значительные изменения потребления кислорода и выделение углекислого газа. Гипоксия и понижение рН крови (ацидоз) вызывают усиление вентиляции (гипервентиляция), а гипероксия и повышение рН крови (алкалоз) - понижение вентиляции (гиповентиляция) или апноэ. Контроль за нормальным содержанием во внутренней среде организма О2, СО2 и рН осуществляется периферическими и центральными хеморецепторами. При гипоксии не только не происходит увеличения глубины дыхания, но даже возникает тенденция к ее уменьшению. Вместе с тем ритм дыхательных движений резко учащается. Этот эффект обусловлен различными причинами. Во-первых, при недостатке кислорода во вдыхаемом воздухе падает содержание СО2 в альвеолярном воздухе. При уменьшении же углекислоты в крови возбуждение дыхательного центра, вызывающее вдох, достигает меньшей силы, чем в нормальных условиях. Во-вторых, дыхательный центр в условиях кислородного голодания становится особенно легко тормозимым. Уже относительно небольшая сила импульсов, проводимых по блуждающим нервам (при относительно малой экскурсии грудной клетки), оказывается достаточной, чтобы прекратить начавшееся возбуждение. В силу этого дыхание делается и более частым, и более поверхностным. Вместе с тем, гипоксия вызывает рефлекторное учащение дыхания, действуя на хеморецепторы каротидного синуса и дуги аорты. Таким образом, эффект кислородного голодания оказывается в достаточной мере сложным. Как правило, гиперкапния производит преимущественно углубление дыхания, гипоксия же вызывает значительное учащение ритма. Наибольшее возрастание объема легочной вентиляции происходит, когда дыхательный центр подвергается одновременному воздействию избытка углекислоты и недостатка кислорода, так как в этом случае происходит одновременно возрастание и глубины, и ритма дыхательных движений.
Так, например, в одном из опытов при вдыхании углекислоты объем вентиляции составлял 81 л/мин при ритме дыхания 30 в минуту и глубине одного дыхания, равной 2,7 л. Когда при том же содержании углекислоты во вдыхаемом воздухе было уменьшено парциальное напряжение кислорода, объем вентиляции возрос до 131 л, причем ритм участился до 66 в минуту, а глубина дыхания уменьшилась до 1,98 л. Наибольшие величины легочной вентиляции наблюдаются у человека во время тяжелой мышечной работы, так как в этом случае дыхательный центр подвергается одновременному воздействию гиперкапнии и аноксии. Адекватным раздражителем для периферических хеморецепторов является уменьшение напряжения кислорода артериальной крови, в меньшей степени увеличение напряжения углекислого газа и рН, а для центральных хеморецепторов - увеличение концентрации Н+ во внеклеточной жидкости мозга. Артериальные (периферические) хеморецепторы. Периферические хеморецепторы находятся в каротидных и аортальных тельцах. Сигналы от артериальных хеморецепторов по синокаротидным и аортальным нервам первоначально поступают к нейронам ядра одиночного пучка продолговатого мозга, а затем переключаются на нейроны дыхательного центра. Недостаток кислорода в артериальной крови является основным раздражителем периферических хеморецепторов. Гипоксическая реакция дыхания практически отсутствует у коренных жителей высокогорья и исчезает примерно через 5 лет у жителей равнин после начала их адаптации к высокогорью (3500 м и выше). Окончательно не установлено местоположение центральных хеморецепторов. Исследователи считают, что такие хеморецепторы находятся в ростральных отделах продолговатого мозга вблизи его вентральной поверхности, а также в различных зонах дорсального дыхательного ядра. Наличие центральных хеморецепторов доказывается достаточно просто: после перерезки синокаротидных и аортальных нервов у подопытных животных исчезает чувствительность дыхательного центра к гипоксии, но полностью сохраняется реакция дыхания на гиперкапнию (повышенное содержание углекислого газа) и ацидоз. Перерезка ствола мозга непосредственно выше продолговатого мозга не влияет на характер этой реакции. Адекватным раздражителем для центральных хеморецепторов является изменение концентрации Н+ во внеклеточной жидкости мозга. Функцию регулятора пороговых сдвигов рН в области центральных хеморецепторов выполняют структуры гематоэнцефалического барьера, который отделяет кровь от внеклеточной жидкости мозга. Через этот барьер осуществляется транспорт 02, С02 и Н+ между кровью и внеклеточной жидкостью мозга. Транспорт С02 и Н+ из внутренней среды мозга в плазму крови через структуры гематоэнцефалического барьера регулируется с участием фермента карбоангидразы. Гиперкапния и ацидоз стимулируют, а гипокапния и алкалоз тормозят центральные хеморецепторы. Для определения чувствительности центральных хеморецепторов к изменению рН внеклеточной жидкости мозга используют метод возвратного дыхания. Испытуемый дышит из замкнутой емкости, заполненной предварительно чистым О2. При дыхании в замкнутой системе выдыхаемый СО2 вызывает линейное увеличение концентрации СО2 и одновременно повышает концентрацию Н+ в крови, а также во внеклеточной жидкости мозга. Тест проводят в течение 4-5 мин под контролем содержания СО2 в выдыхаемом воздухе.
|
|||||||||
|
|
Последнее изменение этой страницы: 2017-02-17; просмотров: 322; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.222.134.54 (0.012 с.) |